Amino Acid and Lipid Rhythmicity in the Menstrual Cycle: Metabolic Fluctuations and Research Implications
This article synthesizes current research on the rhythmic fluctuations of amino acids and lipids across the menstrual cycle, a critical area for drug development and precision medicine.
Amino Acid and Lipid Rhythmicity in the Menstrual Cycle: Metabolic Fluctuations and Research Implications
Abstract
This article synthesizes current research on the rhythmic fluctuations of amino acids and lipids across the menstrual cycle, a critical area for drug development and precision medicine. We explore the foundational science behind phase-dependent metabolic shifts, including the significant decrease in amino acids and specific lipid species during the luteal phase. The content details advanced methodological approaches, such as multi-platform metabolomics and lipidomics, for capturing these dynamics. It further addresses key challenges in study design, including participant stratification and phase verification, and evaluates the validation of metabolic biomarkers for clinical applications. Aimed at researchers and drug development professionals, this review provides a comprehensive framework for integrating menstrual cycle metabolic rhythmicity into biomedical research and therapeutic innovation.
The Metabolic Clockwork of the Menstrual Cycle: Foundational Rhythms in Amino Acids and Lipids
The menstrual cycle represents a fundamental biological rhythm characterized by dynamic fluctuations in key hormones, primarily estrogen and progesterone, which collectively govern an extensive regulatory network over metabolic processes. These hormonal shifts create a complex interplay that directly influences macronutrient metabolism, energy homeostasis, and cellular signaling pathways throughout the body. Research demonstrates that these hormonal variations induce rhythmic patterns in metabolites, including amino acids and lipids, across distinct menstrual cycle phases [1]. This metabolic rhythmicity underscores the profound influence of reproductive hormones on physiological processes extending far beyond the reproductive system. Within this framework, estrogen and progesterone emerge as central regulators, modulating metabolic pathways through both complementary and antagonistic mechanisms, with significant implications for women's health, disease susceptibility, and therapeutic development.
Molecular Mechanisms of Estrogen and Progesterone in Metabolic Regulation
Estrogen Signaling Pathways
Estrogen exerts its metabolic effects primarily through genomic and non-genomic signaling mechanisms mediated by estrogen receptors (ERs), including ERα, ERβ, and the membrane-bound GPER. The binding of estrogen to its nuclear receptors initiates transcriptional regulation of genes involved in glucose and lipid metabolism [2]. Additionally, non-genomic signaling occurs rapidly through membrane-associated receptors and involves second messenger systems.
A key metabolic pathway influenced by estrogen is the insulin signaling cascade. Estrogen enhances insulin sensitivity by promoting the phosphorylation of insulin receptor substrate (IRS) proteins and activating the PI3K-Akt pathway [3]. This activation leads to the phosphorylation of Forkhead box O (FoxO) transcription factors, resulting in their cytoplasmic retention and subsequent suppression of hepatic gluconeogenic genes such as phosphoenolpyruvate carboxykinase (PEPCK) and glucose-6-phosphatase (G6Pase) [3]. Furthermore, estrogen signaling converges with insulin signaling on critical regulators such as mTORC1 and Sirt1, creating an integrated network for metabolic control [3].
In lipid homeostasis, estrogen modulates the activity of key enzymes including malonyl-CoA decarboxylase, acetyl-CoA carboxylase, and fatty acid synthase, thereby reducing malonyl-CoA availability and long-chain fatty acid synthesis [2]. This results in decreased de novo lipogenesis, reduced ectopic lipid accumulation in insulin-sensitive tissues, and improved overall insulin sensitivity [2].
Progesterone Signaling and Metabolic Influence
While research has historically focused more extensively on estrogen, progesterone plays a significant and complementary role in metabolic regulation. Progesterone signals through its specific nuclear progesterone receptors (PR-A and PR-B) and membrane-associated receptors to influence metabolic processes. Its rising levels during the luteal phase contribute to the metabolic shifts observed in this cycle phase.
Progesterone appears to have complex interactions with insulin signaling pathways, potentially creating a state of relative insulin resistance during the luteal phase to support potential pregnancy [1]. This may be mediated through cross-talk with glucocorticoid signaling pathways or through modulation of insulin receptor affinity. Additionally, progesterone influences mitochondrial function and energy expenditure, potentially contributing to the observed increases in basal body temperature during the luteal phase.
The combined effects of estrogen and progesterone create a sophisticated regulatory system that maintains metabolic flexibility across the menstrual cycle, adapting energy utilization and substrate metabolism to support potential reproductive demands.
Quantitative Metabolic Changes Across the Menstrual Cycle
Amino Acid and Protein Metabolism
Comprehensive metabolic profiling reveals significant rhythmicity in amino acid levels across menstrual cycle phases, with the most pronounced decreases observed during the luteal phase. This pattern suggests a state of increased protein turnover or utilization during this period.
Table 1: Amino Acid Fluctuations Across Menstrual Cycle Phases
| Metabolite Class | Specific Metabolites | Direction of Change in Luteal Phase | Statistical Significance |
|---|---|---|---|
| Amino Acids & Derivatives | Ornithine, Arginine, Alanine, Glycine, Methionine, Proline | Significant decrease | p < 0.05 in all 5 phase contrasts [1] |
| Biogenic Amines | Multiple species | Significant decrease | 37 metabolites reached FDR <0.20 for L-M contrast [1] |
| Total Glutathione | Glutathione | Significant decrease | p < 0.05 for L-F contrast [1] |
The consistent reduction in amino acid concentrations during the luteal phase may indicate increased protein synthesis or nitrogen utilization to support potential endometrial preparation for implantation [1]. This metabolic adaptation represents a significant shift in substrate utilization that coincides with peak progesterone levels.
Lipid Metabolism and Distribution
Lipid species demonstrate distinct rhythmic patterns across the menstrual cycle, with notable phase-dependent fluctuations in phospholipids and other lipid classes. These changes reflect the complex interplay between estrogen and progesterone in regulating lipid homeostasis.
Table 2: Lipid Profile Changes Across Menstrual Cycle Phases
| Lipid Parameter | Change Direction | Cycle Phase with Most Significant Change | Research Findings |
|---|---|---|---|
| Phospholipids (LPCs, PCs, LPEs) | Significant decrease | Luteal phase | 17 lipid species met FDR <0.20 for L-F contrast [1] |
| Cholesterol Profiles | Non-linear variation | Throughout cycle | Significant association with menstrual cycle phase (p < 0.001) [4] |
| LDL-C, TC, TG | Increase | Late perimenopause & early postmenopause | SWAN study findings [2] |
| HDL-C | Initial increase, then plateau | Late postmenopause | SWAN study findings [2] |
The transition to central adiposity during perimenopause represents a significant shift in fat distribution patterns, moving from the gynoid (femoral-gluteal) pattern characteristic of premenopausal years to a more android pattern associated with increased cardiometabolic risk [2]. This redistribution is strongly influenced by the decline in estrogen levels and contributes to the metabolic changes observed during the menopausal transition.
Experimental Protocols for Menstrual Cycle Metabolic Research
Longitudinal Metabolic Phenotyping
The comprehensive characterization of metabolic rhythmicity across the menstrual cycle requires rigorous experimental design and precise phase determination. The following protocol outlines a standardized approach for longitudinal metabolic phenotyping:
Cycle Phase Classification:
- Menstrual phase (M): Days 1-5 of cycle, characterized by low estrogen and progesterone
- Follicular phase (F): Days 6-12, rising estrogen levels
- Periovulatory phase (O): Days 13-15, estrogen peak followed by LH surge
- Luteal phase (L): Days 16-26, high progesterone and estrogen
- Premenstrual phase (P): Days 27-28, declining hormone levels
Sample Collection and Verification:
- Collect blood, urine, or other biofluids at each phase
- Verify cycle timing using urinary luteinizing hormone (LH) tests
- Confirm hormonal status with serum estradiol and progesterone measurements
- Record menstrual symptoms and cycle characteristics [1]
Metabolomic Analysis:
- Employ multiple analytical platforms including LC-MS and GC-MS for comprehensive coverage
- Analyze amino acids, biogenic amines, lipids, acylcarnitines, and organic acids
- Utilize standardized normalization procedures to account for technical variation
- Apply multivariate statistical analysis to identify phase-specific metabolic patterns [1]
Specialized Method for Minimal-Invasive Sampling
Recent methodological advances enable comprehensive metabolic profiling from minimal sample volumes, facilitating dense longitudinal sampling:
Finger-Prick Blood Collection:
- Collect approximately 100 μL of capillary blood via finger-prick
- Process samples immediately or store at -80°C until analysis
- This approach enables frequent sampling across cycle phases with reduced participant burden [5]
Multi-Platform Metabolomic Analysis:
- Implement five complementary analytical methods to cover diverse metabolite classes
- Quantify steroid hormones, lipid mediators, abundant lipids, and hydrophilic metabolites
- Analyze 692+ metabolites from a single 100 μL sample [5]
Data Integration and Biomarker Identification:
- Apply principal component analysis (PCA) and partial least squares discriminant analysis (PLS-DA) to identify discriminatory metabolites
- Calculate metabolite ratios to enhance biomarker performance
- Validate findings against clinical symptoms and pain scores [5]
Signaling Pathways in Hormonal Metabolic Regulation
Estrogen-Insulin Signaling Crosstalk
The intricate crosstalk between estrogen and insulin signaling pathways forms a critical regulatory network for metabolic homeostasis. The following diagram illustrates the key molecular interactions and convergence points between these systems:
Estrogen-Insulin Signaling Crosstalk Diagram
This integrated signaling network demonstrates how estrogen potentiates insulin action through multiple mechanisms, including enhanced IRS-PI3K-Akt signaling, regulation of FoxO1 transcriptional activity, and modulation of appetite-regulating neurons in the hypothalamus [3]. The convergence on mTORC1 and Sirt1 represents key integration points for nutrient sensing and energy homeostasis.
Central Regulation of Metabolism via Gut-Brain Axis
The gut-brain axis serves as a crucial interface for metabolic regulation, with hormonal signals from the gastrointestinal tract influencing central regulatory centers. The following diagram illustrates the central signaling pathways of key metabolic hormones:
Gut-Brain Axis Signaling Pathways Diagram
These pathways demonstrate how hormonal signals from the gastrointestinal tract, such as GLP-1 and GLP-2, communicate with central regulatory centers to coordinate metabolic responses including appetite regulation, insulin secretion, and glucose homeostasis [6]. The GLP-1 system is particularly significant as it represents a major therapeutic target for metabolic disorders.
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Essential Research Reagents for Menstrual Cycle Metabolic Studies
| Reagent/Material | Specific Examples | Research Application | Key Function |
|---|---|---|---|
| LC-MS/MS Systems | Triple quadrupole MS, High-resolution MS | Metabolite quantification | Simultaneous analysis of multiple metabolite classes with high sensitivity [1] |
| Hormone Assay Kits | Estradiol ELISA, Progesterone RIA, LH Urinalysis | Cycle phase verification | Confirm menstrual cycle phase and hormonal status [1] |
| Metabolomic Standards | Isotope-labeled amino acids, Lipid standards, Steroid mixes | Quantification calibration | Internal standards for precise metabolite quantification [5] |
| Cell Culture Models | Primary hepatocytes, Neuronal cell lines, Adipocyte models | Mechanistic studies | Elucidate tissue-specific hormone actions and signaling pathways [3] |
| Animal Models | Ovariectomized mice, ER knockout models, Perimenopausal models | In vivo validation | Study systemic metabolic effects of hormonal manipulation [2] |
| Molecular Biology Kits | qPCR assays, Western blot antibodies, Chromatin IP kits | Signaling pathway analysis | Detect gene expression, protein phosphorylation, and transcriptional regulation [3] |
This toolkit enables researchers to investigate the complex metabolic effects of estrogen and progesterone across multiple biological scales, from molecular mechanisms to whole-organism physiology. The integration of data from these diverse approaches is essential for advancing our understanding of hormonal regulation in metabolism.
The intricate interplay between estrogen and progesterone in regulating metabolic processes creates a dynamic system that adapts to the changing hormonal landscape of the menstrual cycle. The rhythmicity in amino acid and lipid metabolism underscores the profound influence of these hormones on substrate utilization and energy homeostasis. Understanding these mechanisms provides critical insights for developing targeted interventions for menstrual-related disorders, optimizing metabolic health across the lifespan, and advancing women-specific healthcare strategies. Future research should focus on elucidating the molecular details of progesterone signaling, exploring tissue-specific hormonal effects, and investigating how modifiable factors such as nutrition and physical activity interact with hormonal status to influence metabolic outcomes. The methodological advances in metabolic profiling and minimal-invasive sampling present exciting opportunities for high-resolution mapping of metabolic rhythmicity in diverse populations and physiological states.
Documenting Global Metabolomic Shifts: LC-MS and GC-MS Evidence Across Cycle Phases
The menstrual cycle represents a fundamental biological rhythm that profoundly influences systemic metabolism. Advanced metabolomic techniques, including liquid chromatography-mass spectrometry (LC-MS) and gas chromatography-mass spectrometry (GC-MS), have begun to elucidate the precise molecular oscillations that occur across cycle phases. This technical review synthesizes evidence from pivotal studies that document consistent, significant rhythmicity in amino acid and lipid species, characterized by a pronounced decrease in the luteal phase. These cyclic metabolic patterns, driven by fluctuating estrogen and progesterone levels, create a framework for understanding periodicity in nutrient utilization and vulnerability to hormone-sensitive disorders. Establishing these baseline rhythmic patterns is crucial for developing cycle-informed clinical interventions and nutritional strategies in women's health.
The menstrual cycle is an essential, approximately 28-day physiological rhythm governed by tightly regulated interactions between progesterone, estradiol, follicular stimulating hormone (FSH), and luteinizing hormone (LH) [7] [8]. Beyond its reproductive function, this cycle exerts systemic metabolic influence, creating predictable patterns in nutrient utilization and energy homeostasis. The integration of high-throughput metabolomic technologies has provided unprecedented insight into the scope of these biochemical oscillations, revealing rhythmicity in pathways central to neurotransmitter synthesis, glutathione metabolism, and urea cycling [1].
Understanding these metabolomic shifts is paramount within broader research on amino acid and lipid rhythmicity. The menstrual cycle presents a natural model for studying how hormonal drivers coordinate systemic metabolic changes, with implications for conditions exhibiting catamenial patterns, such as premenstrual dysphoric disorder (PMDD), phenylketonuria (PKU), and other inborn errors of metabolism [9] [10]. This whitepaper details the experimental methodologies, key findings, and biological implications of global metabolomic shifts across the menstrual cycle, providing a technical foundation for researchers and drug development professionals.
Experimental Protocols for Menstrual Cycle Metabolomics
Cohort Design and Sample Collection
Robust experimental design is critical for capturing authentic metabolic fluctuations. Key studies have established standardized protocols for longitudinal sampling in healthy, premenopausal women.
- Participant Selection: Studies typically enroll healthy, premenopausal women (e.g., n=34 [7] [8]) aged 18-45 with self-reported regular menstrual cycles (21-35 days) for at least three prior cycles. Exclusion criteria commonly include hormonal contraceptive use, pregnancy, lactation, smoking, chronic medical conditions, and recent medication use known to affect metabolism or hormonal balance.
- Cycle Phase Classification: Accurate phase classification is achieved through a multi-modal approach combining:
- Self-reported timing: Participant records of menstrual cycle start date and cycle length.
- Serum hormone quantification: Concentrations of progesterone, estradiol, FSH, and LH are measured to biochemically verify cycle phase.
- Urinary luteinizing hormone (LH): Used to pinpoint the periovulatory LH surge. This integrated data allows for precise classification into five distinct phases: Menstrual (M), Follicular (F), Periovulatory (O), Luteal (L), and Premenstrual (P) [7] [8].
- Biofluid Collection: Fasting blood samples are collected during each participant's classified phase. Plasma, serum, and urine are processed according to standard protocols and stored at -80°C until analysis to preserve metabolite integrity.
Metabolomic Profiling Using LC-MS and GC-MS
Comprehensive metabolomic coverage requires complementary analytical platforms. The following workflow is representative of high-quality studies in this field [7] [8] [1]:
- Sample Preparation: Plasma and urine samples are thawed and prepared for analysis. Protein precipitation is typically performed for plasma samples using cold organic solvents (e.g., methanol or acetonitrile) to remove proteins and recover metabolites. The supernatant is then analyzed directly or derivatized.
- Liquid Chromatography-Mass Spectrometry (LC-MS):
- Function: Ideal for analyzing a wide range of semi-polar and polar metabolites, including complex lipids, amino acids, biogenic amines, and vitamins.
- Protocol: Reverse-phase chromatography separates metabolites based on hydrophobicity. The eluent is then introduced into a high-resolution mass spectrometer via electrospray ionization (ESI). This platform is extensively used for lipidomics and targeted analysis of specific metabolite classes [7].
- Gas Chromatography-Mass Spectrometry (GC-MS):
- Function: Best suited for the analysis of volatile, non-polar, or thermally stable metabolites. It is highly effective for profiling organic acids, sugars, and small acids.
- Protocol: Metabolites often require chemical derivatization (e.g., silylation) to increase their volatility and thermal stability. GC separates the derivatized compounds, which are then ionized by electron impact (EI) before mass analysis [8].
- Data Processing and Statistical Analysis: Raw data from LC-MS and GC-MS are processed using software (e.g., Shimadzu Labsolutions, Neonatal Solutions) for peak picking, alignment, and metabolite identification against standard libraries [9]. Subsequent statistical analysis involves:
- Univariate analysis: Paired t-tests or ANOVA to identify significantly altered metabolites between phases (p-value < 0.05).
- Multiple testing correction: False Discovery Rate (FDR) control (e.g., q < 0.20) to minimize Type I errors.
- Multivariate analysis: Principal Component Analysis (PCA) or Partial Least Squares-Discriminant Analysis (PLS-DA) to visualize group separations and identify key discriminatory metabolites [11].
The following diagram illustrates the integrated experimental workflow from participant recruitment to data analysis.
Figure 1: Experimental workflow for menstrual cycle metabolomics studies, integrating longitudinal sampling, multi-modal phase classification, and complementary LC-MS/GC-MS profiling.
Key Metabolomic Findings: Amino Acid and Lipid Rhythmicity
Quantitative Evidence of Amino Acid Fluctuations
Metabolomic analyses consistently reveal significant oscillations in amino acid and biogenic amine concentrations across the menstrual cycle, with the most pronounced decrease occurring during the luteal phase.
Table 1: Significant Amino Acid and Biogenic Amine Changes Across the Menstrual Cycle (Luteal vs. Other Phases)
| Metabolite Class | Example Metabolites | Change in Luteal Phase | Statistical Significance | Key Phase Contrasts |
|---|---|---|---|---|
| Amino Acids & Derivatives | Ornithine, Arginine, Alanine, Glycine, Proline [8] | ↓ Decrease | FDR q < 0.20 [7] | Luteal vs. Menstrual (L-M), Luteal vs. Follicular (L-F) |
| Biogenic Amines | Serine, Methionine, Threonine, Glutamine [8] | ↓ Decrease | p < 0.05 [1] | Luteal vs. Follicular (L-F), Luteal vs. Periovulatory (L-O) |
| Urea Cycle Metabolites | Citrulline [7] | ↓ Decrease | FDR q < 0.20 [7] | Luteal vs. Menstrual (L-M) |
In a comprehensive study of 34 women, 39 amino acids and derivatives showed a significant decrease (FDR < 0.20) in the luteal phase [7] [1]. This pattern suggests a state of increased nitrogen utilization and potential protein anabolism during the period of elevated progesterone [8]. This rhythmicity is not limited to healthy populations; research in patients with classic phenylketonuria (PKU) shows statistically significant fluctuations in phenylalanine (Phe) concentrations (F(2.85, 271.13) = 5.79, p < 0.001), with the lowest levels in the early luteal phase and the highest during the early follicular phase (menstruation) [9] [10].
Lipid Metabolic Rhythmicity
The lipidome also exhibits distinct cyclical patterns, particularly affecting phospholipid species.
Table 2: Significant Lipid Species Changes Across the Menstrual Cycle
| Lipid Class | Example Lipid Species | Change in Luteal Phase | Statistical Significance | Key Phase Contrasts |
|---|---|---|---|---|
| Phospholipids | Lysophosphatidylcholines (LPCs), Phosphatidylcholines (PCs) [1] | ↓ Decrease | FDR q < 0.20 [8] | Luteal vs. Follicular (L-F) |
| Complex Lipids | Lysophosphatidylethanolamine (LPE 22:6) [8] | ↓ Decrease | p < 0.05 | Luteal vs. Follicular (L-F), Luteal vs. Menstrual (L-M) |
Of 139 plasma lipid species analyzed, 57 were significantly altered (p < 0.05), with 38% consistently decreasing in the luteal phase compared to the follicular and menstrual phases [8] [1]. This collective reduction in amino acids and lipids points to a systemic metabolic adaptation, possibly an anabolic state, during the luteal phase, with recovery occurring during menstruation and the follicular phase [7].
Biological Interpretation and Signaling Pathways
The observed metabolomic patterns are not random but reflect a coordinated physiological response to the changing hormonal landscape. The luteal phase, characterized by a peak in progesterone and a secondary rise in estrogen, appears to induce a state of increased metabolic demand and anabolic potential.
- Hormonal Regulation: The inverse relationship between progesterone/estradiol levels and concentrations of amino acids and lipids suggests these hormones directly or indirectly stimulate utilization pathways. For instance, the course of phenylalanine in PKU patients is inverse to the estradiol concentration, suggesting a direct hormonal effect on metabolism [10].
- Anabolic State: The widespread decrease in circulating amino acids and specific lipids in the luteal phase may indicate their increased uptake and incorporation into tissues, a process potentially driven by progesterone [7] [8].
- Energy Substrate Shift: Observations of increased acetoacetate and VLDL levels in the luteal phase suggest a shift in energy metabolism [11]. This may represent an increased reliance on lipid oxidation for energy, sparing glucose and amino acids for other anabolic processes.
The following diagram synthesizes these concepts into a proposed signaling pathway linking hormonal changes to observed metabolomic outcomes.
Figure 2: Proposed biological pathway linking luteal phase hormonal shifts to observed metabolomic changes through increased anabolism and altered energy substrate use.
The Scientist's Toolkit: Essential Research Reagents and Materials
Executing robust menstrual cycle metabolomics requires specific, high-quality materials and reagents. The following table details key solutions used in the featured experiments.
Table 3: Key Research Reagent Solutions for Menstrual Cycle Metabolomics
| Reagent / Material | Function / Application | Example Use Case in Protocol |
|---|---|---|
| LC-MS Grade Solvents (Methanol, Acetonitrile) | Protein precipitation and mobile phase preparation for LC-MS. | Precipitating proteins from plasma samples prior to LC-MS injection [7]. |
| Derivatization Reagents (e.g., MSTFA, BSTFA) | Chemical modification of metabolites for GC-MS analysis. | Silylation of organic acids and other polar metabolites to increase volatility for GC-MS separation [8]. |
| Stable Isotope-Labeled Internal Standards | Normalization for quantitative accuracy in mass spectrometry. | Added to each biofluid sample to correct for variations in sample preparation and instrument performance [9]. |
| Neonatal Screening Kits (e.g., Neomass AAAC Plus) | Standardized extraction of amino acids from dried blood spots (DBS). | Extraction of phenylalanine and other amino acids from patient-collected DBS for PKU monitoring [9]. |
| Hormone Assay Kits (e.g., ELISA, CLIA) | Quantification of serum progesterone, estradiol, LH, and FSH. | Biochemical verification of menstrual cycle phase for accurate sample classification [7] [8]. |
| C18 Reverse-Phase LC Columns | Chromatographic separation of semi-polar and non-polar metabolites. | Separating complex lipid species (e.g., LPCs, PCs) and amino acids prior to MS detection [1]. |
LC-MS and GC-MS evidence unequivocally demonstrates that the menstrual cycle is a key determinant of metabolic rhythmicity in premenopausal women. The documented global shifts, particularly the luteal phase decrease in amino acids and specific lipid species, provide a metabolic signature of a healthy, rhythmic state. This signature serves as a crucial baseline from which to investigate vulnerabilities to hormone-related health issues such as PMS, PMDD, and catamenial exacerbation of inborn errors of metabolism.
Future research must focus on several critical areas. First, there is a need to elucidate the precise molecular mechanisms by which estrogen and progesterone regulate the transporters and enzymes responsible for the observed metabolomic shifts. Second, the translational application of this knowledge is imperative; dietary guidelines and nutritional strategies, particularly for women with metabolic disorders, should be investigated for efficacy when tailored to the menstrual cycle phase [12]. Finally, the integration of metabolomic data with other omics layers (e.g., transcriptomics, proteomics) will provide a systems-level understanding of this fundamental biological rhythm, ultimately paving the way for more personalized and effective healthcare interventions for women.
The menstrual cycle represents a critical physiological rhythm with profound implications for metabolic homeostasis. Emerging evidence indicates that the luteal phase, characterized by elevated progesterone, is a period of significant amino acid depletion, suggesting a systemic anabolic state. This whitepaper synthesizes findings from metabolomic, lipidomic, and clinical studies demonstrating consistent reductions in plasma amino acids, derivatives, and specific lipid species during the luteal phase. These rhythmic fluctuations occur within the context of neurotransmitter precursor metabolism, glutathione metabolism, and urea cycle activity, potentially creating vulnerability to hormone-related disorders while simultaneously preparing the endometrium for potential pregnancy. The comprehensive metabolic patterns outlined provide a foundation for novel nutritional strategies and therapeutic interventions tailored to the unique physiological states of the menstrual cycle.
The menstrual cycle is governed by intricate interactions between progesterone, estradiol, follicular stimulating hormone (FSH), and luteinizing hormone (LH), creating a rhythmic pattern that extends beyond reproductive function to influence systemic metabolism [8] [1]. Within this framework, the luteal phase specifically demonstrates a pronounced metabolic shift characterized by decreased circulating amino acids and altered lipid profiles [8] [1] [13]. This paper examines the evidence supporting the hypothesis that these metabolic changes represent a physiological anabolic state, potentially directed toward preparing the endometrium for implantation and supporting early pregnancy.
Research utilizing advanced metabolomic and lipidomic profiling technologies has revealed that of 397 metabolites and micronutrients tested, 208 show significant changes (p < 0.05) throughout the menstrual cycle, with 71 meeting the false discovery rate threshold of 0.20 [8] [1]. These fluctuations exhibit distinct patterns across cycle phases, with the most pronounced amino acid depletion occurring during the luteal phase, coinciding with the progesterone peak [8]. Understanding these rhythmic metabolic patterns provides crucial insights for developing cycle-specific nutritional interventions and pharmaceutical approaches that align with natural female physiology.
Comprehensive Metabolic Fluctuations Across the Menstrual Cycle
Amino Acid and Biogenic Amine Dynamics
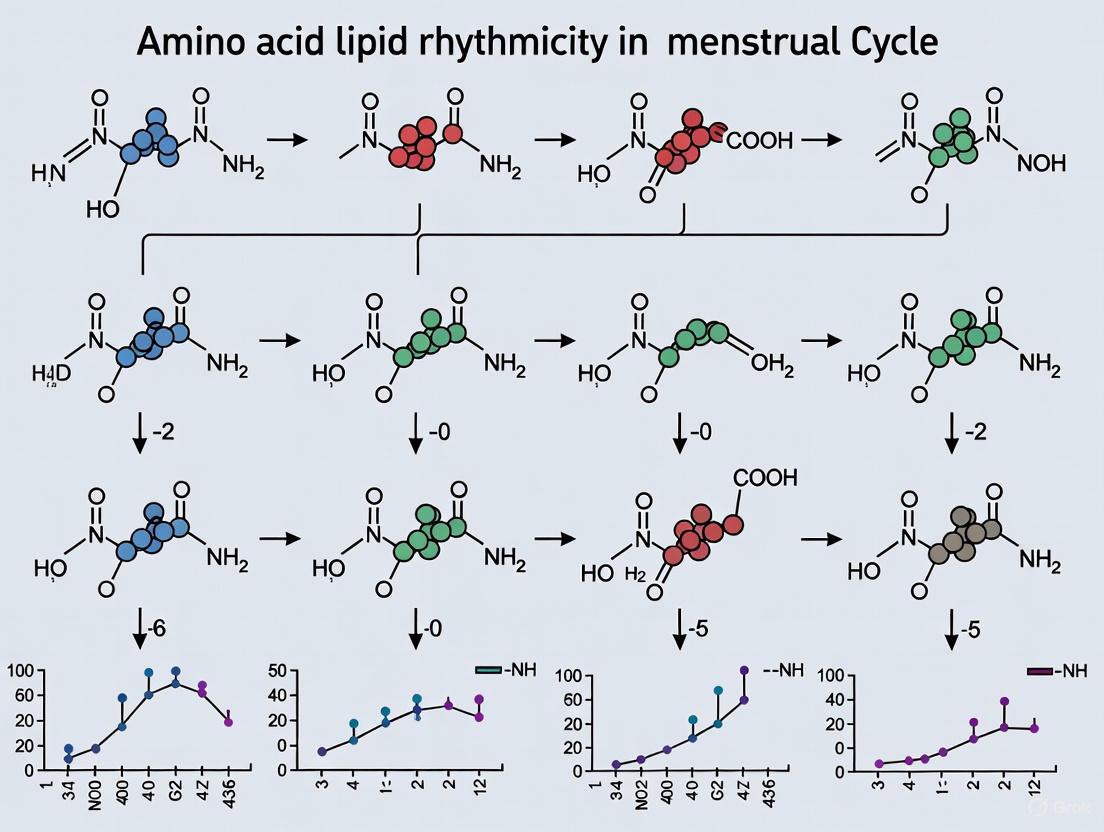
Quantitative analyses reveal significant phase-specific fluctuations in amino acids and biogenic amines, with the most pronounced decreases observed during the luteal phase. A comprehensive study of 54 amino acids and derivatives found that 48 reached statistical significance (p < 0.05) in phase contrast comparisons, with 37 meeting the FDR threshold (q < 0.20) for the luteal-menstrual phase contrast [8] [1].
Table 1: Significant Amino Acid Reductions in Luteal Phase (FDR < 0.20)
| Amino Acid | Luteal-Follicular Effect Size | Luteal-Menstrual Effect Size | Biological Significance |
|---|---|---|---|
| Ornithine | -0.35 | -0.47 | Urea cycle intermediate, polyamine precursor |
| Arginine | -0.34 | -0.47 | Substrate for NO synthase and arginase |
| Threonine | -0.45 | -0.59 | Glycogenic amino acid |
| Alanine | -0.35 | -0.45 | Gluconeogenic substrate |
| Glycine | -0.31 | -0.40 | Glutathione synthesis, neurotransmitter |
| Serine | -0.26 | -0.37 | Phospholipid synthesis |
| Methionine | -0.25 | -0.37 | Methyl group donor, glutathione precursor |
The observed decreases in amino acids appear to be progesterone-associated. A dedicated study investigating arginine dynamics found a significant decline at the luteal phase directly correlated with progesterone levels [13]. This progesterone-associated arginine decrease was accompanied by related fluctuations in ornithine and citrulline, suggesting coordinated regulation of the urea cycle and nitric oxide pathways [13].
Lipid Species Variations
Lipid profiling demonstrates parallel decreases in specific lipid classes during the luteal phase. Of 139 lipid species with detectable plasma levels, 57 reached statistical significance (p < 0.05), with 17 meeting the FDR threshold (q < 0.20) for the luteal-follicular contrast [8] [1].
Table 2: Lipid Species Significantly Reduced in Luteal Phase
| Lipid Class | Representative Species | Phase Contrast | Potential Functional Role |
|---|---|---|---|
| Lysophosphatidylcholines (LPC) | LPC 16:0, LPC 18:0, LPC 18:1 | L-F, L-M | Inflammatory modulation, membrane precursors |
| Phosphatidylcholines (PC) | PC aa 34:2, PC aa 36:2, PC aa 38:4 | L-F, L-M | Membrane composition, signaling precursors |
| Lysophosphatidylethanolamines (LPE) | LPE 22:6 | L-F, L-M, L-O, P-L | Endocannabinoid system modulation |
The simultaneous reduction of both amino acids and phospholipids during the luteal phase suggests a coordinated metabolic shift rather than isolated biochemical phenomena. This pattern may indicate increased utilization of these substrates for tissue preparation and immune modulation [8] [13].
Mechanisms Underlying Luteal Phase Metabolic Changes
Proposed Physiological Workflow
The metabolic adaptations observed during the luteal phase represent a coordinated physiological response to changing hormonal milieus. The following diagram illustrates the proposed workflow through which progesterone elevation drives amino acid depletion and creates an anabolic state:
Molecular and Cellular Mechanisms
The metabolic shifts observed during the luteal phase are driven by several interconnected biological mechanisms:
Protein Turnover and Nitrogen Metabolism: The luteal phase demonstrates increased whole-body protein turnover, evidenced by elevated nitrogen excretion and amino acid oxidation [13]. This enhanced protein flux provides substrates for potential tissue anabolism, particularly endometrial preparation. The observed decrease in plasma amino acids may reflect increased cellular uptake and utilization for protein synthesis rather than catabolic processes.
Urea Cycle and Immune Modulation: Significant decreases in arginine, ornithine, and citrulline suggest coordinated activation of urea cycle pathways [8] [13]. Arginine serves as substrate for both arginase (producing ornithine and urea) and nitric oxide synthase (producing NO and citrulline). Increased arginase activity during the luteal phase may support polyamine synthesis through ornithine conversion while simultaneously reducing arginine availability for T-cell receptor CD3ζ expression, potentially contributing to the immune tolerance necessary for potential embryo implantation [13].
Glutathione and Oxidative Stress Management: Cyclical changes in glutathione metabolism components suggest rhythmic regulation of antioxidant defenses [8]. The luteal phase demonstrates alterations in glutamate, glycine, and cysteine availability - all glutathione precursors. This coordinated adjustment may represent preparation for managing oxidative stress associated with endometrial remodeling.
Experimental Approaches and Methodologies
Comprehensive Metabolomic Workflow
The evidence supporting luteal phase amino acid depletion derives from sophisticated experimental approaches integrating multiple analytical platforms. The following diagram illustrates a representative workflow for comprehensive metabolic profiling across menstrual cycle phases:
Detailed Methodological Protocols
Participant Selection and Cycle Phase Classification: Studies typically enroll healthy, premenopausal women with regular menstrual cycles (28 ± 7 days) and no hormonal contraceptive use [8] [1] [13]. Phase classification employs multiple verification methods including serum hormone measurements (progesterone, estradiol, FSH, LH), urinary luteinizing hormone detection, and basal body temperature tracking [8] [13]. The five-phase classification system includes menstrual (M), follicular (F), periovulatory (O), luteal (L), and premenstrual (P) phases [8].
Sample Collection and Processing: Blood collection occurs in the morning after fasting to minimize diurnal variation effects [13]. Studies utilize various sample types including plasma (for metabolomics, lipidomics, amino acids), serum (for clinical chemistries and hormones), and urine (for metabolic profiling) [8] [1]. For self-sampling protocols, participants collect capillary blood via finger-prick onto filter paper for dried blood spot analysis [14].
Analytical Platforms:
- Liquid Chromatography-Mass Spectrometry (LC-MS): Employed for targeted and untargeted analysis of amino acids, biogenic amines, and lipid species [8] [1]
- Gas Chromatography-Mass Spectrometry (GC-MS): Used for organic acid profiling and additional metabolomic coverage [8]
- High-Performance Liquid Chromatography with Fluorescence Detection (HPLC-FLD): Applied for precise quantification of B vitamins and related cofactors [8]
- Immunoassays: Utilized for hormone quantification (progesterone, estradiol, FSH, LH) to verify cycle phases [8] [13]
Statistical Analysis: Data analysis incorporates mixed-effects models to account for repeated measures within participants [8]. Significance thresholds typically employ p < 0.05 with false discovery rate (FDR) correction (q < 0.20) for multiple testing [8] [1]. Multivariate analyses including principal component analysis (PCA) and partial least squares discriminant analysis (PLS-DA) identify metabolic patterns differentiating cycle phases [5].
Research Reagent Solutions Toolkit
Table 3: Essential Research Materials and Analytical Platforms
| Category | Specific Reagents/Platforms | Research Application | Key Features |
|---|---|---|---|
| Hormonal Assays | Serum progesterone & estradiol immunoassays | Cycle phase verification | Quantifies primary cycle regulators |
| Metabolomics | LC-MS platforms with targeted panels | Amino acid & amine quantification | High sensitivity for polar metabolites |
| Lipidomics | GC-MS & LC-MS lipid profiling | Phospholipid & acylcarnitine analysis | Broad coverage of lipid classes |
| Sample Collection | Dried blood spot filter cards | Self-sampling & stability | Enables frequent home collection |
| Statistical Analysis | R/Python with mixed-effects models | Longitudinal data analysis | Accounts for within-subject variation |
Clinical Implications and Research Applications
The rhythmic amino acid depletion observed during the luteal phase has significant implications for both clinical practice and pharmaceutical development. In women with inborn errors of metabolism such as phenylketonuria (PKU), the menstrual cycle significantly influences metabolic control, with the lowest phenylalanine concentrations occurring in the early luteal phase and the highest during the early follicular phase [14] [9]. This pattern appears independent of dietary intake, suggesting direct hormonal effects on amino acid metabolism [9].
For drug development, these cyclic metabolic patterns suggest potential phase-dependent differences in drug metabolism and amino acid-derived therapeutic efficacy. The consistent decrease in specific amino acid classes during the luteal phase may influence the pharmacokinetics of amino acid-based therapeutics or drugs metabolized through amino acid-dependent pathways.
From a nutritional science perspective, these findings raise important questions about potential phase-specific protein and amino acid requirements. While some early studies suggested increased protein oxidation during the luteal phase, recent evidence indicates that protein requirements do not change substantially across the menstrual cycle [15]. However, the observed metabolic rhythmicity may still inform personalized nutrition strategies for women with specific metabolic conditions or performance goals.
The comprehensive metabolic profiling of menstrual cycle phases provides compelling evidence that the luteal phase represents a distinct physiological state characterized by coordinated amino acid depletion and specific lipid profile alterations. These changes support the hypothesis of a progesterone-driven anabolic state potentially directed toward endometrial preparation and immune modulation. The consistent patterns observed across multiple studies and analytical platforms underscore the importance of considering menstrual cycle phase as a critical variable in female metabolism research.
Future research directions should include mechanistic studies to elucidate the molecular pathways connecting progesterone signaling to amino acid transport and utilization, investigation of cycle-phase effects on drug metabolism and therapeutic efficacy, and clinical trials exploring cycle-specific nutritional interventions for women with metabolic disorders. Integrating these metabolic rhythms into the broader context of female physiology will enable more precise and effective healthcare strategies tailored to women's unique physiological patterns.
The menstrual cycle represents a fundamental biological rhythm, driven by cyclical fluctuations in key hormones such as estradiol, progesterone, follicular stimulating hormone (FSH), and luteinizing hormone (LH). These hormonal changes regulate not only reproduction but also exert a significant influence on systemic metabolism. A growing body of evidence indicates that lipid metabolism is particularly sensitive to these rhythmic hormonal shifts. Understanding the fluctuations of specific lipid classes—triglycerides, wax esters, and phospholipids—across the menstrual cycle provides crucial insights into female physiology and forms an essential component of a broader thesis investigating the interplay between amino acid and lipid rhythmicity. This cyclical metabolic pattern may underlie susceptibility to hormone-related health issues and opens avenues for novel nutritional and therapeutic strategies tailored to cyclic physiology [1]. This whitepaper synthesizes current research to serve as a technical guide for researchers, scientists, and drug development professionals working in metabolic health and women's medicine.
Comprehensive metabolic profiling studies have revealed distinct rhythmic patterns for various lipid classes throughout the menstrual cycle. The data, derived from advanced analytical techniques including LC-MS and GC-MS, are summarized in the following tables for clear comparison.
Table 1: Phospholipid Fluctuations Across Menstrual Cycle Phases
| Phospholipid Species | Observed Fluctuation Pattern | Significant Phase Contrasts (q < 0.20) | Proposed Physiological Implication |
|---|---|---|---|
| Total Phospholipids | Significant decrease in the luteal phase [1] | Luteal vs. Follicular (L-F), Luteal vs. Menstrual (L-M) [1] | Membrane remodeling, altered lipid signaling |
| LPC Species | 6 LPC species significantly reduced in luteal phase [1] | L-F contrast [1] | Inflammation modulation, energy metabolism |
| PC Species | 10 PC species significantly reduced in luteal phase [1] | L-F contrast [1] | Cell membrane integrity, one-carbon metabolism |
| LPE 22:6 | Statistically significant difference in 4 out of 5 phase contrasts [1] | L-F, L-M, L-O, P-L [1] | Docosahexaenoic acid (DHA) metabolism, neuro-endocrine signaling |
Table 2: Wax Esters and Fatty Acid Composition in Meibum
| Analyte | Observed Fluctuation Pattern | Key Phases of Change | Association/Implication |
|---|---|---|---|
| Total Wax Esters (WE) | Cyclic change in fatty acid (FA) composition of WEs [16] | Phases II & III (Mid-Cycle) vs. other phases [16] | Alters meibum viscosity & melting temperature [16] |
| Saturated FAs (e.g., C16:0, C18:0) | Positive correlation with PC1 (77.5%) in PCA [16] | Higher in Phases II & III cluster [16] | Increases melting temperature |
| Monounsaturated FAs (e.g., C18:1n9) | Negative correlation with PC1 in PCA [16] | Higher in cluster excluding Phases II & III [16] | Decreases melting temperature, maintains fluidity |
Note on Triglycerides (TG): The search results indicate that triglyceride levels did not demonstrate a consistent, statistically significant rhythmic variation across the menstrual cycle in several studies [17] [1]. However, targeted metabolomics in specific pathologies like PCOS have shown elevated triglycerides, particularly those containing unsaturated fatty acids [18].
Detailed Experimental Protocols and Methodologies
To ensure reproducibility and critical evaluation of the data, this section outlines the key methodological approaches used in the cited research.
Metabolomic and Lipidomic Profiling in Serum/Plasma
This protocol is adapted from studies that identified phospholipid rhythmicity [1].
- 1. Sample Collection: Blood samples are collected from healthy, premenopausal women at multiple, precisely timed points across a confirmed ovulatory menstrual cycle. Key phases include Menstrual (M), Follicular (F), Periovulatory (O), Luteal (L), and Premenstrual (P).
- 2. Sample Preparation: Plasma or serum is separated via centrifugation. For lipidomics, proteins are precipitated using cold methanol. Lipids are then extracted using a biphasic solvent system, such as a mixture of methyl-tert-butyl ether (MTBE)/methanol/water, which separates lipids into an organic phase [1] [5].
- 3. Lipid Analysis:
- LC-MS/MS for Phospholipids: The extracted lipidome is separated using reverse-phase or hydrophilic interaction liquid chromatography (HILIC) and analyzed with a high-resolution mass spectrometer (e.g., Q-TOF or Orbitrap) [1].
- Data Processing: Raw data is processed using specialized software for peak picking, alignment, and identification by matching against lipid databases (e.g., LIPID MAPS).
- 4. Statistical Analysis: Data is normalized and log-transformed. Linear mixed models are used to account for repeated measures. Significance is tested for phase-phase contrasts (e.g., Luteal vs. Follicular), with False Discovery Rate (FDR) correction for multiple testing (e.g., q < 0.20 considered significant) [1].
Analysis of Wax Esters and Fatty Acids in Meibum
This protocol is derived from the study analyzing meibum composition [16].
- 1. Subject Classification & Cycle Phase Determination: Healthy premenopausal women with regular cycles are recruited. The menstrual cycle is divided into phases (e.g., I-VI) based on the first day of menstruation and basal body temperature (BBT) charting to confirm ovulation.
- 2. Meibum Collection: After a warm compress, meibum is non-invasively collected from the meibomian gland orifices using a sterilized Daviel spoon or similar tool, avoiding contamination from tears or skin lipids.
- 3. Lipid Extraction and Derivatization: The meibum sample is dissolved in an organic solvent (e.g., chloroform/methanol). An internal standard is added. Fatty acids are hydrolyzed from complex lipids (like wax esters and cholesteryl esters) and derivatized to fatty acid methyl esters (FAMEs) using a reagent like boron trifluoride-methanol.
- 4. Gas Chromatography-Mass Spectrometry (GC-MS): The FAME derivatives are separated by gas chromatography and detected/identified by mass spectrometry. The relative abundance of each fatty acid is quantified by comparing peak areas to the internal standard.
- 5. Data Analysis: Principal Component Analysis (PCA) is performed on the fatty acid composition data to identify clustering patterns associated with menstrual cycle phases [16].
Signaling Pathways and Metabolic Workflows
The rhythmic fluctuations of lipids are governed by the interplay of hormones and metabolic pathways. The following diagram illustrates the proposed integrative pathway connecting hormonal shifts to the observed lipid changes.
Integrative Hormonal Regulation of Lipid Rhythmicity. This diagram illustrates the proposed mechanistic pathway. Hormonal fluctuations drive the rhythmicity by modulating key metabolic enzymes and processes. Estrogen rise stimulates lipoprotein lipase and LCAT activity, influencing phospholipid remodeling and LDL/HDL cholesterol profiles [17]. Concurrently, hormonal shifts modulate fatty acid synthesis and sebaceous gland secretion, leading to changes in the fatty acid composition of wax esters in tissues like meibum [16]. The observed decrease in many plasma phospholipids and amino acids in the luteal phase may indicate a state of increased anabolic utilization [1].
The Scientist's Toolkit: Essential Research Reagents and Materials
This table details key reagents, materials, and instruments essential for conducting research in menstrual cycle lipidomics, as derived from the methodologies cited.
Table 3: Research Reagent Solutions for Menstrual Cycle Lipidomics
| Item/Category | Specific Examples | Function & Application in Research |
|---|---|---|
| Sample Collection & Preparation | EDTA or Heparin tubes (plasma), Serum Separator Tubes, Sterile Daviel spoon (meibum) [16], Organic solvents (Methanol, MTBE, Chloroform) [1] [16] | Biological fluid collection; protein precipitation and lipid extraction from plasma, serum, or meibum. |
| Internal Standards & Reagents | Deuterated lipid standards (e.g., d₇-PC, d₅-TG), Supelco 37 FAME Mix [16], Boron trifluoride-methanol solution [16] | Quantification of specific lipid classes via mass spectrometry; derivatization of fatty acids for GC-MS analysis. |
| Chromatography & Separation | LC-MS grade solvents (Acetonitrile, Isopropanol, Water), HILIC or C18 LC columns [1] [5], GC capillary columns [16] | Separation of complex lipid mixtures prior to mass spectrometric detection. |
| Mass Spectrometry Systems | High-resolution LC-MS/MS (e.g., Q-TOF, Orbitrap) [1], Gas Chromatograph-Mass Spectrometer (GC-MS) [16] | Identification and quantification of thousands of lipid species and metabolites. |
| Data Analysis Software | LCModel (for MRS data) [18], Lipidomics-specific software (e.g., LipidSearch, MS-DIAL), R or Python with statistical packages | Spectral quantification, lipid identification, peak alignment, and statistical analysis for rhythmicity. |
| Hormonal Assay Kits | ELISA or LC-MS/MS kits for Estradiol, Progesterone, LH, FSH [16] [19] | Objective verification of menstrual cycle phase for accurate participant stratification. |
The evidence clearly demonstrates that the menstrual cycle is a significant biological variable influencing lipid metabolism, with phospholipids and wax esters showing distinct phase-dependent fluctuations. These changes are likely mediated by rhythmic hormonal signals and have implications for cellular membrane integrity, lipid signaling, and the function of specialized secretions like meibum. The integration of these lipid dynamics with concurrent rhythmicity in amino acid metabolism, as observed in the simultaneous decrease of both metabolite classes during the luteal phase, presents a compelling systems physiology perspective [1]. Future research should focus on elucidating the precise molecular mechanisms linking hormonal receptors to lipid metabolic enzymes, exploring the impact of these fluctuations on drug efficacy and pharmacokinetics, and developing cycle-phase-specific interventions for metabolic disorders and women's health conditions. Standardizing methodologies and increasing the temporal resolution of sampling in large cohorts will be crucial to fully decode the complex rhythmicity of the lipidome.
Linking Metabolic Rhythmicity to Physiological Demands and Endometrial Biosynthesis
Emerging research reveals that the menstrual cycle is governed by intricate metabolic rhythmicity, extending far beyond canonical sex hormone fluctuations to encompass periodic changes in amino acid, lipid, and energy metabolism. This review synthesizes recent metabolomic and lipidomic evidence characterizing these cyclical patterns in healthy women, framing them within a physiological context of shifting endometrial biosynthetic demands. We provide a detailed analysis of quantitative metabolite variations across cycle phases, delineate the molecular signaling pathways coordinating these rhythms, and present standardized experimental protocols for their investigation. For researchers and drug development professionals, this work underscores the critical importance of accounting for metabolic rhythmicity in experimental design, diagnostic development, and therapeutic targeting of women's health disorders, positioning the menstrual cycle as a central model for understanding metabolic periodicity.
The human menstrual cycle represents a complex, rhythmically oscillating system essential for reproductive fitness and overall metabolic health. While traditionally characterized by its endocrinological profile, advanced metabolomic technologies now reveal that the cycle is underpinned by profound, systemic oscillations in biochemical pathways. These include rhythmic variations in amino acid availability, phospholipid composition, and energy substrate utilization that are tightly coupled to the phases of endometrial proliferation, differentiation, and shedding [1]. This metabolic rhythmicity represents a fundamental adaptation to meet the distinct physiological demands of tissue remodeling, with the endometrium undergoing cycles of rapid growth, secretory transformation, and breakdown. Understanding these patterns is paramount for developing targeted interventions for menstrual-related disorders, optimizing nutritional strategies, and advancing precision medicine approaches that account for cyclical physiological states. This technical guide synthesizes current evidence on metabolic rhythmicity across the menstrual cycle, with a specific focus on its implications for endometrial biosynthesis and systemic metabolic demands.
Quantitative Metabolic Profiles Across the Menstrual Cycle
Comprehensive metabolic profiling studies have systematically quantified rhythmic variations across menstrual cycle phases. The following tables summarize key metabolite changes observed in healthy, premenopausal women, providing a reference for researchers investigating cycle-specific metabolic states.
Table 1: Plasma Amino Acids and Biogenic Amines Showing Significant Phase-Dependent Variations
| Metabolite Class | Specific Metabolites | Direction of Change in Luteal Phase | Significant Contrasts (q < 0.20) | Proposed Physiological Relevance |
|---|---|---|---|---|
| Amino Acids & Derivatives | Ornithine, Arginine, Alanine, Glycine, Methionine, Proline | ↓ Decrease | L-F, L-M, L-O, P-L, O-M (all 5 contrasts) | Reduced availability may indicate increased anabolic demand for protein synthesis [1] |
| Threonine, Ornithine, Serine | ↓ Decrease | L-M, L-F, L-O, P-L (4 contrasts) | ||
| Total Amines Significantly Altered | 37 compounds | ↓ Decrease | L-M contrast | Potential vulnerability period for hormone-related health issues [1] |
| Urea Cycle Components | Ornithine, Arginine | ↓ Decrease | Consistent across phases | Implication of urea cycle rhythmicity in nitrogen handling [1] |
Table 2: Lipid Species and Micronutrients Showing Significant Phase-Dependent Variations
| Analyte Category | Specific Compounds | Direction of Change | Significant Phase Contrasts | Proposed Physiological Relevance |
|---|---|---|---|---|
| Phospholipids | LPE 22:6 | ↓ Decrease in Luteal | L-F, L-M, L-O, P-L | Membrane remodeling and prostaglandin precursor availability [1] |
| 6 LPCs, 10 PCs, 1 LPE | ↓ Decrease in Luteal | L-F (q < 0.20) | ||
| Micronutrients | 25-OH Vitamin D | ↑ Increase in Menstrual | L-M, O-M (q < 0.20) | Potential cyclical regulation of immune and calcium metabolism [1] |
| Pyridoxic Acid (Vitamin B6 metabolite) | ↑ Increase in Menstrual | O-M (q < 0.20) | Altered vitamin B6 status potentially affecting neurotransmitter synthesis [1] | |
| Energy Metabolism | Glucose | ↓ Decrease in Luteal | L-M, L-O, L-P (p < 0.05) | Cyclical energy substrate preference and insulin sensitivity [1] |
The data presented in these tables originate from a study of 34 healthy premenopausal women, with biofluids collected at multiple timepoints and analyzed via LC-MS, GC-MS, and HPLC-FLD, followed by statistical analysis with false discovery rate (FDR) correction [1]. A key finding is the widespread decrease in plasma amino acids and specific lipid species during the luteal phase, potentially indicative of an anabolic state during the progesterone peak, with recovery occurring during menstruation and the follicular phase [1]. These systematic fluctuations provide a quantitative foundation for understanding the metabolic basis of cyclic physiological demands.
Experimental Methodologies for Investigating Metabolic Rhythmicity
Robust investigation of menstrual cycle metabolic rhythmicity requires stringent experimental design, precise phase determination, and appropriate statistical analysis.
Subject Recruitment and Cycle Phase Classification
- Participant Criteria: Recruit healthy, premenopausal women (e.g., age 18-45) with self-reported regular menstrual cycles (25-35 days) and not using hormonal contraception. Exclude participants with diagnosed endocrine disorders, PCOS, or recent pregnancy/lactation [1].
- Phase Determination: Utilize a multi-modal classification system combining:
- Serum Hormone Measurements: Quantify estradiol, progesterone, FSH, and LH via immunoassays.
- Urinary Luteinizing Hormone: Track LH surge to pinpoint ovulation.
- Self-Reported Timing: Record first day of menses and cycle length.
- Five-Phase Classification: Define samples according to five distinct phases [1]:
- Menstrual (M): Days 1-5 of cycle.
- Follicular (F): Post-menstruation, pre-ovulatory.
- Periovulatory (O): Within 24-48 hours of detected LH surge.
- Luteal (L): Post-ovulation, characterized by high progesterone.
- Premenstrual (P): Late luteal phase, following progesterone peak.
Metabolomic and Lipidomic Profiling Protocols
- Sample Collection & Preparation:
- Collect plasma (EDTA/K2EDTA tubes), serum, and 24-hour urine samples at each phase.
- Immediately process samples: centrifuge (e.g., 2000 × g, 10 min, 4°C), aliquot, and store at -80°C.
- For LC-MS/MS analysis, precipitate proteins with cold methanol or acetonitrile followed by centrifugation and supernatant analysis.
- Instrumental Analysis:
- Broad-Spectrum Metabolomics: Employ untargeted LC-MS (Reversed-Phase and HILIC) and GC-MS to maximize metabolite coverage [1].
- Targeted Lipidomics: Use multiple reaction monitoring (MRM) on a triple quadrupole mass spectrometer for precise quantification of lipid species (e.g., LPCs, PCs, LPEs) [1].
- Micronutrient Analysis: Quantify B vitamins and metabolites like pyridoxic acid using HPLC with fluorescence detection (HPLC-FLD) [1].
- Quality Control:
- Incorporate pooled quality control (QC) samples from all study samples.
- Use internal standards for metabolite classes (e.g., stable isotope-labeled amino acids) to correct for instrument variation.
Statistical Analysis and Data Interpretation
- Data Preprocessing: Normalize data to account for urine dilution (e.g., using creatinine) and correct for batch effects.
- Statistical Testing:
- Employ linear mixed-effects models to account for repeated measures within subjects.
- Test for significant differences (p < 0.05) in metabolite levels across predefined phase contrasts (e.g., Luteal vs. Follicular: L-F).
- Apply False Discovery Rate (FDR) correction for multiple testing (e.g., q < 0.20 threshold) to control for false positives [1].
- Advanced Analytical Considerations:
- For high-dimensional data (e.g., >1000 metabolites in nontargeted studies), sparse multivariate methods like sparse partial least squares (SPLS) are recommended, especially with larger sample sizes, as they demonstrate greater selectivity and lower potential for spurious relationships compared to univariate methods [20].
- Consider using relative changes (e.g., fold-change) or metrics like Canberra distance that are less biased by absolute concentration differences between metabolites [21].
Molecular Mechanisms Linking Metabolism and Endometrial Biosynthesis
The observed systemic metabolic rhythms are not merely correlative but are mechanistically intertwined with the endometrial cycle through specific molecular pathways.
Hormonal Regulation of Nutrient Partitioning
- Progesterone-Driven Anabolism: The pronounced decrease in plasma amino acids and lipids during the luteal phase coincides with the progesterone peak, suggesting a hormone-mediated redirection of nutrients toward endometrial protein synthesis and membrane lipid incorporation in preparation for potential implantation [1] [22].
- Nitrogen Utilization Rhythmicity: Nitrogen balance studies reveal a statistically significant (p < .001) biphasic cycle in urinary nitrogen excretion in women, implying intrinsic hormonal regulation of nitrogen metabolism that is unique to females and independent of protein intake [22].
Metabolic Reprogramming in Endometrial Tissue
In pathological states like endometriosis, ectopic endometrial cells exhibit cancer-like metabolic reprogramming, including a Warburg-like effect (aerobic glycolysis) even in normoxic conditions [23]. This reprogramming, driven by pathways such as PI3K/AKT/mTOR and HIF-1α signaling, ensures a constant supply of energy and biosynthetic precursors (e.g., amino acids, nucleotides, lipids) for lesion survival and growth [23]. While exaggerated in disease, this highlights the inherent metabolic plasticity of endometrial tissue, which is likely subject to more subtle, rhythmic regulation during the normal cycle to support periodic biosynthesis.
Inflammatory and Immune Coordination
The luteal phase is accompanied by a state of low-grade inflammation, which, in a healthy cycle, is precisely regulated. Cytokines such as IL-6 and TNF-α can directly interfere with insulin and sex hormone receptor signaling in the endometrium [24] [25]. This crosstalk creates a nexus where inflammatory signals, nutrient availability, and hormonal status converge to coordinate endometrial remodeling, with breakdown occurring if implantation does not ensue.
This table details key reagents, assays, and software essential for conducting research in menstrual cycle metabolic rhythmicity.
Table 3: Essential Research Reagents and Resources
| Item Name / Category | Specific Examples / Formats | Primary Function in Research Context |
|---|---|---|
| Immunoassay Kits | Estradiol, Progesterone, LH, FSH ELISA/Chemiluminescence Kits | Precise hormonal quantification for accurate menstrual cycle phase classification [1]. |
| Mass Spectrometry Standards | Stable Isotope-Labeled Amino Acid Mixes, Lipid Internal Standards (e.g., d7-Cholesterol), ({}^{15})N-Glycine | Internal standardization for accurate quantification of metabolites in complex biological matrices via LC-MS/GC-MS [1] [22]. |
| Chromatography Columns | C18 (Reversed-Phase), HILIC, GC capillary columns | Separation of diverse metabolite classes (lipids [C18], polar compounds [HILIC]) prior to mass spectrometric detection [1]. |
| Cell Signaling Assays | Phospho-specific antibodies for AKT, mTOR; HIF-1α ELISA Kits | Investigation of molecular pathways (PI3K/AKT, HIF) driving metabolic reprogramming in endometrial cells [24] [23]. |
| Statistical & Bioinformatics Software | R packages (lme4 for mixed models), Python (Scikit-learn), CalR (for energy expenditure), Pathway Analysis Tools (MetaboAnalyst) | Handling high-dimensional data, performing complex repeated-measures statistics, and conducting pathway enrichment analysis [20] [26]. |
The integration of metabolomics with gynecologic research has definitively established that the menstrual cycle is a phenomenon of systemic metabolic rhythmicity, characterized by predictable oscillations in amino acids, lipids, and micronutrients. These rhythms are not merely incidental but are functionally linked to the profound biosynthetic demands of the cyclic endometrium. For the research and drug development community, these findings carry significant implications: experimental design must account for cycle phase as a critical biological variable, diagnostic biomarkers for conditions like PCOS and endometriosis may reside in their characteristic disruption of these metabolic rhythms, and therapeutic strategies could be optimized by timing interventions to specific metabolic phases. Future research must leverage advanced multivariate statistical models [20] and multi-omics integration to further elucidate the genetic and molecular regulators of this rhythmicity, ultimately paving the way for cycle-aware precision medicine in women's health.
From Sampling to Systems Biology: Methodologies for Capturing Metabolic Rhythmicity
The integration of lipidomics, metabolomics, and hormone assays represents a transformative approach in women's health research, particularly for investigating the complex rhythmic interactions between amino acids and lipids across the menstrual cycle. This multi-omics paradigm enables the systematic mapping of metabolic fluctuations driven by hormonal changes, revealing novel biomarker patterns associated with gynecological conditions such as endometriosis and dysmenorrhea. Advanced mass spectrometry techniques, coupled with machine learning analytics, are uncovering dynamic metabolic signatures that promise to revolutionize diagnostics, therapeutic monitoring, and personalized treatment strategies. This technical guide provides researchers and drug development professionals with comprehensive methodologies, experimental workflows, and current findings essential for implementing these integrated platforms in menstrual cycle research.
The menstrual cycle represents one of the fundamental biological rhythms in human physiology, governed by complex interactions between reproductive hormones and metabolic pathways. Research has established that approximately 70% of plasma metabolites are lipids, making lipidomics a particularly valuable tool for understanding metabolic changes throughout the cycle [27]. When combined with metabolomic profiling of amino acids, hormones, and other small molecules, researchers can obtain a comprehensive view of the biochemical changes that occur across menstrual phases. This integrated approach is especially powerful for investigating conditions like endometriosis, where multi-omics analyses have demonstrated significantly improved diagnostic performance over single-platform methodologies [28]. The emerging field of amino acid lipid rhythmicity specifically focuses on how these metabolite classes fluctuate in concert with hormonal changes, creating unique metabolic patterns that may serve as biomarkers for both physiological and pathological states.
Menstrual Cycle Rhythmicity: Key Metabolic Findings
Amino Acid and Lipid Fluctuations Across Cycle Phases
Comprehensive metabolic profiling has revealed significant rhythmicity in amino acid and lipid levels across physiologically distinct menstrual phases. A foundational study analyzing 397 metabolites and micronutrients found that 208 showed significant changes (p < 0.05) across the cycle, with 71 meeting the false discovery rate threshold of 0.20 [1]. These fluctuations follow distinct patterns that reflect the underlying hormonal regulation of metabolism throughout the menstrual cycle.
Table 1: Amino Acid Fluctuations Across Menstrual Cycle Phases
| Metabolite Class | Specific Metabolites | Direction of Change | Phase of Maximum Change |
|---|---|---|---|
| Amino Acids & Derivatives | Ornithine, arginine, alanine, glycine, methionine, proline | Significant decrease | Luteal phase [1] |
| Total Amines | 37 compounds | Significant decrease | Luteal vs. Menstrual [1] |
| Branched-Chain Amino Acids (BCAAs) | Valine, leucine, isoleucine | Decrease ~0.8-fold | Menstrual phase (pain association) [5] |
| Phospholipids | 6 LPCs, 10 PCs, 1 LPE | Significant decrease | Luteal phase [1] |
| Phosphatidylinositols | PI(36:2) | 1.42-fold increase | Menstrual phase (pain association) [5] |
The decrease in amino acids and derivatives during the luteal phase may indicate an anabolic state during the progesterone peak, with recovery occurring during menstruation and the follicular phase [1]. This pattern suggests increased protein synthesis or nitrogen utilization when progesterone levels are elevated. Similarly, the reduction in phospholipid species during the luteal phase points to cycle-specific reorganization of lipid metabolism, potentially related to endometrial preparation for potential implantation.
Biomarker Potential for Gynecological Conditions
The rhythmic fluctuations of amino acids and lipids across the menstrual cycle not only reflect physiological changes but also show distinctive alterations in gynecological disorders, providing valuable biomarker potential.
Table 2: Multi-Omics Biomarkers in Gynecological Conditions
| Condition | Sample Type | Key Metabolite Alterations | Diagnostic Performance |
|---|---|---|---|
| Endometriosis | Plasma & Peritoneal Fluid | 20 metabolites in PF, 26 in plasma; PC ae C30:2 consistently increased [28] [29] | AUC 0.84 (PF), 0.95 (plasma) - luteal phase [28] |
| Menstrual Pain Severity | Plasma | PI(36:2) increased; BCAAs decreased; PI(36:2)/BCAA ratio [5] | AUC 0.990 for ratio [5] |
| Heavy Menstrual Bleeding | Genetic & longitudinal data | Genetic variants, environmental factors, symptoms [30] | Research ongoing [30] |
For endometriosis diagnosis, a multi-omics approach combining metabolomic and proteomic data has demonstrated superior classification performance compared to either method alone, with sensitivity and specificity reaching 0.98 and 0.86 respectively for plasma assays [28]. Similarly, for objective assessment of menstrual pain severity, the ratio of phosphatidylinositol PI(36:2) to branched-chain amino acids has shown remarkable discrimination power with AUC values of 0.990, significantly outperforming individual metabolites [5].
Analytical Methodologies and Experimental Protocols
Integrated Multi-Omics Workflow
A standardized workflow for integrating lipidomics, metabolomics, and hormone assays is essential for generating reproducible, high-quality data in menstrual cycle research. The following diagram illustrates a comprehensive multi-omics workflow:
Sample Collection and Preparation Protocols
Sample Collection Considerations:
- Timing and Phase Determination: Precise menstrual cycle phase determination is critical. Methods include last menstrual period tracking, urinary luteinizing hormone (LH) tests for ovulation detection, hormonal measurements (serum progesterone, estradiol), and wearable device data (skin temperature, heart rate) [31].
- Sample Types: Plasma, peritoneal fluid, sebum, and urine each provide complementary metabolic information. For endometriosis research, simultaneous collection of plasma and peritoneal fluid is recommended [28] [29].
- Standardized Processing: Immediate processing (<45 minutes) and centrifugation protocols (e.g., 2,500 × g for 10 minutes at 4°C for plasma) are essential for sample integrity [28]. For sebum analysis, standardized skin cleaning and Sebutape patch application for 30 minutes provide reproducible results [32].
Metabolomic Sample Preparation (Plasma/Peritoneal Fluid):
- Thaw samples on ice and centrifuge at 2,750 g, 4°C for 5 minutes
- Follow AbsoluteIDQ p180 kit instructions: add 10 µl internal standard to 96-well plate
- Add 10 µl sample to designated wells, dry under nitrogen stream for 30 minutes
- Derivatize with 50 µl derivatization mix (25 minutes at room temperature)
- Dry for 60 minutes, then add 300 µl extraction solvent
- Vortex (450 RPM, 30 minutes), centrifuge (500 g, 2 minutes)
- Transfer eluted samples to LC-MS and FIA-MS plates for analysis [28]
Lipidomics Sample Preparation (Sebum):
- Extract sebum lipids from Sebutape patches using ethanol with 0.025% BHT
- Perform clean-up with ethyl acetate
- Dissolve extract in acetone/methanol/isopropanol (40/40/20 v/v/v)
- Prepare aliquots in amber glass vials, evaporate to dryness
- Store at -80°C until analysis
- For analysis, reconstitute in AMI mixture [32]
Instrumental Analysis Techniques
Lipidomics Platforms:
- Untargeted Lipidomics: Utilizing high-resolution mass spectrometry (HRMS) such as Q-TOF, Orbitrap, or Fourier transform ion cyclotron resonance MS for comprehensive lipid profiling. Data-independent acquisition (DIA) modes enable broad biomarker discovery [27].
- Targeted Lipidomics: Employing UPLC-triple quadrupole MS with multiple reaction monitoring (MRM) for precise quantification of specific lipid classes. This approach offers superior sensitivity for low-abundance lipids [27].
- GC-MS Analysis: For quantification of squalene, cholesterol, free fatty acids, and fatty alcohols after derivatization with BSTFA + 1% TCMS [32].
Metabolomics Platforms:
- LC-MS/MS with FIA-MS/MS: Using kits such as AbsoluteIDQ p180 enables simultaneous quantification of 188 metabolites including amino acids, biogenic amines, acylcarnitines, glycerophospholipids, and sphingolipids [28].
- Hydrophilic Metabolite Profiling: Complementary LC-MS methods optimized for polar metabolites including branched-chain amino acids, 4-hydroxyproline, and other water-soluble biomarkers [5].
Hormone Assays:
- Conventional Immunoassays: ELISA for steroid hormones (progesterone, estradiol, 17-hydroxyprogesterone) and gonadotropins (LH, FSH) for cycle phase confirmation.
- MS-Based Hormone Profiling: LC-MS/MS panels for simultaneous quantification of 23 steroid hormones with higher specificity and dynamic range [5].
Computational Integration and Data Analysis
Machine Learning for Phase Identification and Biomarker Discovery
Advanced computational methods are essential for extracting meaningful biological insights from multi-omics datasets. Machine learning algorithms can identify subtle patterns in complex data that traditional statistical methods might miss.
Menstrual Phase Classification Using Wearable Data: Random forest models applied to physiological signals from wrist-worn devices (skin temperature, electrodermal activity, interbeat interval, heart rate) can classify menstrual phases with 87% accuracy for three-phase classification (period, ovulation, luteal) with AUC-ROC of 0.96 [31]. This approach reduces participant burden while providing objective phase identification crucial for timing sample collection.
Biomarker Discovery Workflows:
- Data Preprocessing: Normalization, missing value imputation (e.g., 0.5*LOQ for values below quantification limit), and quality control [28]
- Multivariate Analysis: Principal component analysis (PCA) and partial least squares-discriminant analysis (PLS-DA) to identify group separations and influential variables [5]
- Feature Selection: Identification of significantly altered metabolites (fold change >1.2, p<0.015) with false discovery rate correction [5]
- Classification Modeling: Receiver operating characteristic (ROC) analysis to evaluate diagnostic performance of individual metabolites and ratios [5]
- Multi-Omics Integration: Combining metabolomic, lipidomic, and proteomic features to build enhanced classification models [28]
The following diagram illustrates the computational analysis pipeline:
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Essential Research Reagents and Platforms for Integrated Menstrual Cycle Research
| Category | Specific Products/Platforms | Key Applications | Technical Notes |
|---|---|---|---|
| Metabolomics Kits | AbsoluteIDQ p180 Kit (Biocrates) | Quantitative analysis of 188 metabolites | Covers amino acids, biogenic amines, lipids, acylcarnitines [28] |
| Lipidomics Standards | Deuterated internal standards (d6-Cholesterol, d6-Squalene, d5-TG 48:0) | Quantitative lipidomics | Essential for accurate quantification in complex matrices [32] |
| Sample Collection | Sebutape patches (Cuderm S100) | Sebum collection for lipidomics | 30-minute application, standardized surface area [32] |
| Chromatography | UPLC systems (Waters BEH C18 columns) | Compound separation | Essential for resolving complex lipid and metabolite mixtures [28] |
| Mass Spectrometry | Triple quadrupole (QQQ), Q-TOF, Orbitrap platforms | Targeted and untargeted analysis | QQQ for quantification, Q-TOF/Orbitrap for discovery [28] [27] |
| Wearable Sensors | E4 wristband, EmbracePlus, Oura Ring | Physiological monitoring for phase identification | Measures skin temperature, EDA, IBI, HR [31] |
| Data Analysis | R/Python, MetaboAnalyst, MassLynx, TargetLynx | Data processing and statistical analysis | Multivariate statistics, machine learning implementation [28] [5] |
The integration of lipidomics, metabolomics, and hormone assays provides unprecedented insights into the complex metabolic rhythmicity of the menstrual cycle. This multi-omics approach has already demonstrated significant potential for improving diagnosis of gynecological conditions, with biomarker panels showing superior performance compared to single-analyte approaches. The emerging understanding of amino acid-lipid rhythmicity opens new avenues for personalized interventions and chronotherapeutic approaches in women's health.
Future developments in this field will likely focus on several key areas: (1) standardization of multi-omics protocols across research centers to enable larger collaborative studies; (2) integration of wearable device data for real-time metabolic and physiological monitoring; (3) application of artificial intelligence for pattern recognition in complex temporal data; and (4) translation of biomarker discoveries into clinically validated diagnostic tests. As these technologies become more accessible and analytical pipelines more refined, multi-omics approaches promise to transform our understanding of menstrual cycle physiology and pathology, ultimately leading to improved healthcare outcomes for women across the lifespan.
The human menstrual cycle represents a quintessential model of physiological rhythmicity, governed by a complex interplay of endocrine, autocrine, and paracrine factors. For researchers investigating cyclic fluctuations in metabolites such as amino acids and lipids, a rigorous study design for temporal mapping is paramount. The endometrial tissue, the primary uterine site of implantation, undergoes dynamic, cyclical changes of shedding, regeneration, and differentiation throughout reproductive life, processes that are coordinated by the hypothalamic-pituitary-ovarian axis [33]. These structural and functional changes are driven by fluctuating levels of estrogen and progesterone, which not only remodel tissue but also regulate underlying metabolic processes. The first half (follicular phase) is characterized by rising estrogen levels leading to endometrial proliferation, while the second half (luteal phase) is dominated by progesterone, which induces secretory changes and decidualization [33] [8]. Within this context, the study of amino acid and lipid rhythmicity requires precise phase classification and longitudinal sampling strategies to capture biologically meaningful fluctuations that might otherwise be obscured by methodological inconsistency.
Menstrual Cycle Phase Classification Systems
Accurate phase classification is the cornerstone of temporal mapping. The following systems provide frameworks for defining cycle phases with the precision required for metabolic research.
Five-Phase Classification System
A detailed 5-phase system provides the temporal resolution necessary for dense metabolic sampling [8]:
- Menstrual Phase (M): Characterized by low levels of estrogen and progesterone, marking the onset of the cycle.
- Follicular Phase (F): Estrogen levels rise, promoting endometrial regeneration and proliferation.
- Periovulatory Phase (O): Defined by the peak and surge of luteinizing hormone (LH) and follicular stimulating hormone (FSH), culminating in ovulation.
- Luteal Phase (L): Features high levels of both estrogen and progesterone, inducing secretory changes.
- Premenstrual Phase (P): Hormone levels decline, preceding the next menstrual bleed.
Three-Phase Ovarian Cycle Model
For studies where extreme granularity is not required, a simplified 3-phase model is often employed, focusing on ovarian function [34]:
- Follicular (Days 1-10): Low estrogen and progesterone.
- Ovulatory (Days 11-14): High estrogen, low progesterone.
- Luteal (Days 15-28): High estrogen and progesterone.
Table 1: Comparison of Menstrual Cycle Phase Classification Systems
| System | Phases | Key Hormonal Features | Best Use Cases |
|---|---|---|---|
| 5-Phase | Menstrual, Follicular, Periovulatory, Luteal, Premenstrual | Detailed tracking of LH/FSH peaks and hormonal declines | High-density longitudinal sampling; metabolomics studies requiring fine temporal resolution |
| 3-Phase | Follicular, Ovulatory, Luteal | Broad shifts in estrogen and progesterone profiles | Larger cohort studies; longitudinal imaging or functional tests where frequent sampling is impractical |
Longitudinal Sampling Methodologies
Longitudinal sampling designs must balance temporal resolution with practical constraints. The following methodologies, derived from recent studies, provide a framework for effective data collection.
Sampling Protocols for Metabolic Phenotyping
Metabolic profiling requires dense sampling to capture rhythmicity. One protocol involved biofluid collection (plasma and urine) at four timepoints aligned to the 5-phase classification system (Menstrual, Follicular, Periovulatory, Luteal) from 34 healthy, premenopausal women over a single cycle [8]. This design successfully identified significant rhythmicity in 71 metabolites, including amino acids and lipid species. Another study on exhaled volatiles employed a six-interval follow-up schedule, starting from a menstruation baseline and continuing through the subsequent bleed, to monitor volatile organic compound (VOC) concentrations [35]. This high-frequency sampling was critical for identifying pronounced, substance-specific changes, such as a significant decrease in plasma amino acids during the luteal phase [8].
Phase Verification and Biomarker Correlation
Relying solely on self-reported cycle timing is insufficient for rigorous research. Phase classification must be biochemically verified. The following biomarkers are essential for phase confirmation:
- Serum Hormones: Progesterone, estradiol, LH, and FSH levels must be measured to confirm phase classification [8] [36].
- Urinary Luteinizing Hormone: Used as a surrogate marker to pinpoint the LH surge and ovulation [8].
- Machine Learning Augmentation: For studies in free-living conditions, machine learning models incorporating features like "heart rate at the circadian rhythm nadir (minHR)" can improve luteal phase classification and ovulation prediction, outperforming traditional basal body temperature (BBT) methods, especially in individuals with variable sleep patterns [36].
The experimental workflow for establishing a longitudinal sampling design integrates these elements, from participant screening to final analysis.
Analytical Techniques and Data Acquisition
The choice of analytical platform is critical for capturing the full spectrum of metabolic changes.
Metabolomics and Lipidomics Profiling
Advanced mass spectrometry techniques form the backbone of metabolic rhythmicity studies:
- Liquid Chromatography-Mass Spectrometry (LC-MS) and Gas Chromatography-Mass Spectrometry (GC-MS): These are the workhorses for broad-spectrum metabolomics and lipidomics, allowing for the identification and quantification of hundreds of metabolites simultaneously [8].
- Capillary Electrophoresis Time-of-Flight Mass Spectrometry (CE-TOF-MS): This technique is particularly effective for profiling charged metabolites, such as amino acids and their derivatives, and has been used to investigate associations in large cohorts of midlife women [37].
- Proton Transfer Reaction-Time of Flight-MS (PTR-ToF-MS): For real-time, high-resolution monitoring of volatile organic compounds (VOCs) in exhaled breath, this technology enables split-second observation of metabolic changes, providing a non-invasive window into physiology [35].
Single-Cell and Spatial Transcriptomics
To understand the tissue context of metabolic changes, high-resolution molecular techniques are employed:
- Single-Cell RNA Sequencing (scRNA-seq) and Single-Nucleus RNA Sequencing (snRNA-seq): These technologies generate dense cellular maps of tissues like the endometrium, resolving heterogeneity and identifying distinct cell states across the menstrual cycle [33].
- Spatial Transcriptomics (e.g., 10x Genomics Visium): This method maps the spatial location of cell types and gene expression within the tissue architecture, which is crucial for understanding microenvironment-specific signaling [33].
- Computational Integration: Tools like the cell2location algorithm are used to integrate single-cell transcriptomics with spatial data, precisely mapping cell states to specific tissue layers (e.g., lumenal vs. glandular epithelium) [33].
Table 2: Key Analytical Techniques for Temporal Mapping Studies
| Analytical Technique | Measured Analytes | Key Application in Menstrual Cycle Research |
|---|---|---|
| LC-MS / GC-MS | Amino acids, lipids, organic acids | Broad profiling of plasma/urine metabolome; identification of phase-specific metabolic signatures [8] |
| CE-TOF-MS | Charged metabolites (amino acids, urea cycle intermediates) | Targeted analysis of metabolic pathways relevant to hormone-related health issues [37] |
| PTR-ToF-MS | Exhaled Volatile Organic Compounds (VOCs) | Non-invasive, real-time metabolic monitoring; assessment of protein and energy metabolism [35] |
| scRNA-seq / snRNA-seq | Transcriptomes of individual cells | Deconstruction of cellular heterogeneity and identification of hormone-responsive cell states [33] |
| Spatial Transcriptomics | Genome-wide RNA within tissue context | Mapping spatiotemporal dynamics of cell signaling and differentiation in intact tissue [33] |
Experimental Protocols for Core Investigations
Protocol: Metabolomic Profiling of Amino Acids and Lipids
This protocol is adapted from longitudinal studies investigating metabolic rhythmicity [8] [37].
- Participant Preparation: Participants should fast overnight and provide samples at a standardized time of day to control for circadian effects.
- Blood Collection: Collect venous blood into EDTA-containing tubes. Centrifuge at 4°C to separate plasma, which should be aliquoted and stored at -80°C.
- Urine Collection: Collect mid-stream urine. Aliquot after centrifugation and store at -80°C.
- Metabolite Extraction: For plasma, use a methanol-based protein precipitation. For urine, dilute with a solvent compatible with the analytical platform.
- Instrumental Analysis:
- Analyze samples using LC-MS with reverse-phase chromatography for lipid separation and HILIC chromatography for polar metabolites like amino acids.
- Analyze derivatized extracts by GC-MS for a broader profile of organic acids and sugars.
- Data Processing: Use proprietary and open-source software (e.g., XCMS, MS-DIAL) for peak picking, alignment, and compound identification against standard libraries.
Protocol: Establishing 3D Endometrial Organoid Cultures
Endometrial organoids provide a powerful in vitro platform for investigating the mechanisms regulating endometrial differentiation and hormone response [33].
- Tissue Acquisition: Obtain endometrial biopsies from live donors or from uterine tissue post-hysterectomy. Samples should be collected with ethical approval and patient consent.
- Tissue Dissociation: Mechanically mince the tissue and digest with collagenase to dissociate the epithelial glands from the stromal compartment.
- Organoid Culture: Embed the epithelial fragments in a basement membrane matrix (e.g., Matrigel) and culture in a specialized medium containing growth factors (e.g., Wnt agonists, R-spondin, Noggin) and hormones (estrogen, progesterone).
- Hormonal Treatment: To induce differentiation, treat organoids with a physiological sequence of estrogen (to simulate the proliferative phase) followed by progesterone (to simulate the secretory phase).
- Pathway Modulation: To dissect signaling mechanisms, treat organoids with pathway modulators. For example, downregulation of the WNT pathway increases secretory lineage differentiation, while NOTCH pathway inhibition promotes ciliated cell differentiation [33].
- Validation: Characterize organoid response using scRNA-seq to benchmark against in vivo cell states and immunofluorescence for lineage markers (e.g., PAEP for secretory cells, PIFO for ciliated cells).
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Research Reagents for Menstrual Cycle Metabolomics and Tissue Mapping
| Reagent / Material | Function / Application | Example Use Case |
|---|---|---|
| EDTA Plasma Tubes | Stabilizes blood samples for metabolomic analysis by inhibiting coagulation and preserving metabolite integrity. | Collection of plasma for LC-MS/GC-MS analysis of amino acids and lipids [8]. |
| Matrigel | Basement membrane extract providing a 3D scaffold for organoid growth. | Culture of human endometrial epithelial organoids to study hormone response in vitro [33]. |
| Recombinant R-spondin | Activator of Wnt/β-catenin signaling, a critical pathway for stem cell maintenance. | Essential component in the culture medium for maintaining endometrial organoids [33]. |
| 17-β Estradiol & Progesterone | Key ovarian steroid hormones used to simulate menstrual cycle phases in vitro. | Differentiation of endometrial organoids to study secretory and ciliated cell fate [33]. |
| Collagenase | Enzyme for tissue dissociation to isolate specific cell populations or glands. | Digestion of endometrial tissue to extract epithelial glands for organoid culture [33]. |
| Antibody Panel (SOX9, PAEP, PIFO) | Markers for cell identification via immunofluorescence. SOX9: progenitor; PAEP: secretory; PIFO: ciliated. | Validation of cell states in endometrial tissue sections and organoids across cycle phases [33]. |
| WNT/NOTCH Pathway Inhibitors | Chemical modulators to dissect the role of specific signaling pathways in cell differentiation. | Functional testing of pathway regulation on epithelial lineage fate in organoids [33]. |
Data Analysis and Integration
The complex, multi-modal data generated from temporal mapping studies requires sophisticated analytical approaches. Longitudinal Statistical Models must account for the participant-specific nature of repeated measures data. Methods include linear mixed-effects models to test for phase-phase differences, with significance thresholds adjusted for multiple testing using False Discovery Rate (FDR) control [8]. Furthermore, Computational Deconvolution tools are used to interpret bulk data from complex tissues. Cellular maps generated by scRNA-seq can be used as references to deconvolute bulk transcriptomic data from endometrial pathologies, illuminating the cell types dominating in disorders like endometriosis and endometrial carcinoma [33]. Finally, Cell-Cell Communication Analysis is performed using tools like CellPhoneDB v.3.0, which measures intercellular communication by integrating ligand-receptor pair expression with spatial coordinates of cells, defining signaling in specific microenvironments like lumenal and glandular epithelium [33]. The integration of these data types provides a systems-level view of menstrual cycle rhythmicity.
The integration of minimally invasive sampling techniques into modern biomedical research has revolutionized the approach to physiological monitoring, particularly in studies requiring frequent or remote sample collection. Finger-prick blood microsampling—the collection of small volumes of capillary blood (typically ≤ 100 μL) from a fingertip—represents a paradigm shift from conventional venipuncture [38] [39] [40]. This technical guide explores the foundational principles, methodologies, and applications of finger-prick blood analysis, with specific focus on its growing utility in investigating the rhythmic fluctuations of amino acids and lipids across the menstrual cycle. For researchers investigating dynamic metabolic processes such as menstrual cycle rhythmicity, this approach enables dense longitudinal sampling that was previously logistically prohibitive, thereby revealing previously uncharacterized temporal patterns in metabolic profiles [1] [5] [40].
Technical Foundations of Finger-Prick Blood Collection
Core Principles and Advantages
Finger-prick microsampling leverages capillary blood collection via a lancet device to obtain volumes sufficient for sophisticated multi-omic analyses [39]. The technique is characterized by several key advantages:
- Minimal Invasiveness: Causes significantly less pain and stress than venous draws, improving participant compliance, especially for frequent sampling protocols [38] [39].
- Accessibility and Convenience: Enables at-home collection by participants themselves, eliminating geographical and temporal barriers to clinic visits [38] [39] [40].
- High-Quality Molecular Data: When properly collected and stored, capillary blood yields data comparable to venous blood for thousands of metabolites, proteins, and lipids [5] [40].
Recent studies validate that capillary blood concentrations of key biomarkers like Neurofilament Light (NfL) and Glial Fibrillary Acidic Protein (GFAP) are equivalent to those measured in venous serum and plasma, confirming analytical reliability [38].
Essential Materials and Equipment
Table 1: Essential Research Reagent Solutions for Finger-Prick Blood Collection and Analysis
| Item | Function | Specification Considerations |
|---|---|---|
| Sterile Lancets | To perform a quick, small puncture on the fingertip. | Adjustable depth settings are beneficial for varying skin types [41]. |
| Microsampling Devices (e.g., Mitra device) | To collect and stabilize a precise volume of capillary blood. | Based on absorptive microsampling tips; collects ~10-30 μL [39] [40]. |
| Microtainer Tubes (SST for serum, LiHep for plasma) | To collect liquid capillary blood for processing into serum or plasma. | Typically hold 400-600 μL [38]. |
| Alcohol Wipes | To clean the puncture site and prevent sample contamination. | Isopropyl alcohol-based [41]. |
| Transport and Storage Materials | To maintain sample integrity during shipment and storage. | Temperature monitoring is critical for analyte stability [38] [40]. |
Methodological Workflow for Reliable Microsampling
The following experimental protocol synthesizes best practices from recent clinical and research studies [38] [41] [5].
Pre-Collection Preparation
- Subject Preparation: Instruct participants to sit comfortably and warm their hands if cold to promote blood flow [41].
- Site Selection: Typically, the side of the third or fourth finger of the non-dominant hand is preferred due to reduced nerve endings and sufficient capillary density [41].
- Cleaning: Thoroughly clean the selected site with an alcohol wipe and allow it to air dry completely to prevent hemolysis and contamination [41].
Blood Collection and Processing
- Puncture: Use a new, sterile lancet to perform a quick, firm puncture on the side of the fingertip [41].
- Collection: Gently milk the finger to form a blood drop. Wipe away the first drop. For liquid samples, gently collect blood into the appropriate Microtainer tube. For dried blood spots, touch the microsampling device's absorbent tip to subsequent drops until filled [38] [41].
- Crucial Consideration: Avoid excessive "milking" of the finger, as this can dilute the blood with interstitial fluid and compromise analytical results [41].
- Processing: For plasma/serum, process microtainers according to the manufacturer's instructions, often involving centrifugation. For dried samples, allow them to dry completely at room temperature before packaging [38].
Sample Storage and Stability
Analyte stability varies. For instance, NfL remains stable in serum after a seven-day processing delay at ambient temperatures, while many metabolites and lipids are also highly stable [38] [40]. However, stability must be validated for specific analytes.
Application in Menstrual Cycle Metabolic Rhythmicity Research
Finger-prick microsampling is uniquely suited for investigating the complex, temporal metabolic changes that occur throughout the menstrual cycle.
Revealing Cyclic Metabolic Patterns
A landmark study using traditional venous sampling revealed significant rhythmicity in 208 metabolites across the menstrual cycle in healthy women, including decreases in 39 amino acids and derivatives and 18 lipid species during the luteal phase, suggesting a hormone-driven anabolic state [1]. Finger-prick sampling makes profiling these nuanced, intra-individual fluctuations more feasible.
Table 2: Key Metabolites with Rhythmic Patterns Across the Menstrual Cycle
| Metabolite Class | Change During Menstrual Cycle | Potential Physiological Significance |
|---|---|---|
| Amino Acids (e.g., Ornithine, Arginine, Alanine) | Significant decrease in the luteal phase vs. menstrual and follicular phases [1]. | May indicate increased protein/anabolic utilization during the progesterone peak [1]. |
| Phospholipids (e.g., LPCs, PCs, LPE 22:6) | Significant reduction in the luteal phase [1]. | Cyclic remodeling of cell membranes or energy substrate shifts. |
| Branched-Chain Amino Acids (BCAAs) | Lower levels associated with more severe menstrual pain [5]. | Potential marker for inflammatory pain sensitivity. |
| Vitamin D (25-OH) | Higher levels in the menstrual phase [1]. | Possible interaction with hormonal regulation. |
| Total Cholesterol (TC) & LDL-C | Significantly higher in the follicular phase compared to the luteal phase [42] [4]. | Influenced by fluctuating estrogen levels, relevant for cardiovascular risk assessment [42]. |
Advanced Analytical Approaches
The power of microsampling is fully realized when coupled with multi-omic analytical technologies. A proof-of-concept study demonstrated the ability to measure 1,461 metabolites, 128 proteins, and 776 lipids from a single 10-microliter finger-prick sample using mass spectrometry-based platforms [40]. This comprehensive profiling allows researchers to move beyond single biomarkers and capture system-wide metabolic interactions in response to hormonal cycles.
Experimental Protocol for Menstrual Cycle Research
The following provides a detailed methodology for a study investigating amino acid and lipid rhythmicity, based on validated approaches [1] [5] [40].
Study Design and Sampling Schedule
- Participants: Recruit healthy, premenopausal women with confirmed regular ovulatory cycles. Exclude users of hormonal contraception and those with conditions affecting metabolic health.
- Cycle Phase Determination: Use a combination of self-reported cycle timing, urinary luteinizing hormone (LH) tests to pinpoint ovulation, and/or basal body temperature tracking to classify cycle phases accurately (e.g., menstrual, follicular, periovulatory, luteal) [1] [19].
- Sampling Frequency: To capture dynamic changes, collect finger-prick samples at multiple time points within a single cycle. Protocols may include daily sampling during key transitions or 3-5 samples phased around menstruation, ovulation, and the mid-luteal phase [1] [40].
Sample Collection and Analysis
- At-Home Collection: Provide participants with pre-prepared kits containing lancets, microsampling devices (e.g., Mitra), detailed instructions, and pre-labeled packaging.
- Standardized Timing: Instruct participants to collect samples at a consistent time of day (e.g., morning, fasted) to minimize diurnal variation.
- Shipment: Participants mail samples to the central lab via regular post. Studies show many analytes remain stable under ambient shipping conditions for several days [38] [40].
- Laboratory Processing:
- Extraction: Extract molecules from the microsamples using optimized protocols for proteins, lipids, and metabolites [5] [40].
- Multi-Omic Analysis: Employ platforms like Liquid Chromatography-Mass Spectrometry (LC-MS) for metabolomics/lipidomics and immunoassays for specific proteins or hormones [1] [5].
- Data Integration: Correlate the resulting metabolic profiles with the determined menstrual cycle phase and hormone levels to identify rhythmic patterns.
Finger-prick blood microsampling has emerged as a robust, patient-centric methodology that is transforming longitudinal physiological research. Its application to the study of the menstrual cycle effectively addresses long-standing challenges in capturing high-resolution, intra-individual metabolic data. The technique has already proven instrumental in documenting significant rhythmicity in amino acids and lipids, revealing a state of increased anabolic activity during the luteal phase [1] and identifying potential biomarkers for associated symptoms like menstrual pain [5]. As multi-omic technologies continue to advance, the integration of minimally invasive sampling will be crucial for developing a deeper, systems-level understanding of female physiology and for creating personalized health interventions based on cyclic metabolic patterns.
Data Integration and Multi-Omics Strategies for Pathway Analysis
The study of complex biological systems requires a holistic approach that moves beyond single-layer analysis. Data integration and multi-omics strategies for pathway analysis enable researchers to build comprehensive models of biological activity by combining datasets from different molecular levels. When applied to the study of amino acid lipid rhythmicity within the menstrual cycle, these approaches can reveal the intricate metabolic oscillations governed by hormonal fluctuations and their relationship to female physiology and health disorders. This technical guide provides researchers and drug development professionals with advanced methodologies for conducting integrated multi-omics analyses, with specific application to metabolic rhythmicity across the menstrual cycle.
The menstrual cycle represents a dynamic physiological system characterized by rhythmic variations in hormone levels that regulate numerous metabolic processes. Recent metabolomics studies have revealed that approximately 52% of detected metabolites (208 out of 397 tested) show significant variation across menstrual cycle phases, highlighting the profound influence of cyclical hormonal changes on metabolism [1]. Within this context, integrated omics approaches are particularly valuable for identifying molecular markers associated with menstrual symptoms and disorders, as they can capture complementary information from genomic, proteomic, metabolomic, and lipidomic datasets to build unified pathway models.
Core Principles of Multi-Omics Data Integration
Integration Methodologies
Multi-omics data integration strategies can be broadly categorized into three conceptual approaches:
Sequential Integration: Analyzes omics data in a stepwise manner, where results from one analysis inform the next. This approach is useful for establishing potential causal relationships but may miss complex interactions between molecular layers.
Simultaneous Integration: Processes all omics datasets concurrently using multivariate statistical methods, network modeling, or machine learning algorithms. This approach can capture non-linear relationships and complex interactions but requires substantial computational resources.
Model-Based Integration: Incorporates prior biological knowledge to guide the integration process, often using pathway databases or molecular interaction networks as scaffolds for data integration [43].
The selection of an appropriate integration strategy depends on research objectives, data characteristics, and available computational resources. For studying menstrual cycle rhythmicity, simultaneous integration often provides the most comprehensive insights into coordinated fluctuations across metabolic pathways.
Pathway Enrichment Analysis
Pathway enrichment analysis serves as a statistical framework for determining whether defined biological pathways are over-represented in an omics dataset more than would be expected by chance. This method helps researchers translate extensive lists of significant genes, proteins, or metabolites into mechanistic insights about affected biological processes [44].
Two major analytical paradigms exist for pathway enrichment analysis:
Threshold-based methods (e.g., g:Profiler) operate on predefined gene or metabolite lists that meet specific significance criteria. These methods are ideal when working with clearly defined sets of differentially expressed biomolecules.
Rank-based methods (e.g., Gene Set Enrichment Analysis - GSEA) utilize complete ranked lists of all measured molecules without applying arbitrary significance thresholds. These methods preserve information from molecules with modest but coordinated changes [44] [45].
For menstrual cycle studies, rank-based methods often prove advantageous as they can detect subtle but coordinated rhythmicity across entire metabolic pathways.
Experimental Design for Menstrual Cycle Studies
Longitudinal Sampling Strategies
Menstrual cycle research requires carefully timed sampling across multiple cycle phases to capture rhythmic metabolic patterns. The typical experimental design involves collecting biospecimens at defined phases:
- Menstrual phase (days 1-5): Characterized by low estrogen and progesterone levels
- Follicular phase (days 6-12): Rising estrogen levels
- Periovulatory phase (days 13-16): Estrogen peak followed by LH surge
- Luteal phase (days 17-24): Elevated progesterone and estrogen
- Premenstrual phase (days 25-28): Declining hormone levels [1]
To minimize participant burden while enabling comprehensive metabolic profiling, researchers have developed innovative minimally invasive sampling protocols. Recent advancements allow for comprehensive metabolomic analysis from only 100 μL of finger-prick blood, enabling frequent at-home sampling across cycle phases [5].
Multi-Omics Data Generation
A comprehensive multi-omics approach to menstrual cycle rhythmicity should encompass several analytical domains:
Table 1: Essential Omics Domains for Menstrual Cycle Research
| Omics Domain | Analytical Techniques | Key Targets | Relevance to Menstrual Cycle |
|---|---|---|---|
| Metabolomics | LC-MS, GC-MS | Amino acids, lipids, carbohydrates | Direct reflection of metabolic state; reveals cyclic patterns |
| Lipidomics | LC-MS/MS | Phospholipids, triglycerides, cholesterol esters | Membrane fluidity, signaling precursors |
| Proteomics | LC-MS/MS, RPPA | Signaling proteins, receptors, enzymes | Hormone response effectors |
| Hormone Profiling | ELISA, LC-MS/MS | Estradiol, progesterone, LH, FSH | Cycle phase validation |
| Genomics/Transcriptomics | RNA-Seq, microarrays | Gene expression patterns | Regulatory mechanisms |
This multi-layered analytical approach recently identified branched-chain amino acids (BCAAs) and specific phosphatidylinositols (particularly PI(36:2)) as potential biomarkers for menstrual pain severity, with their ratios showing high accuracy in discriminating between mild and severe pain groups (AUC: 0.990 for PI(36:2)/BCAA) [5].
Data Integration Workflows
Preprocessing and Quality Control
Effective data integration begins with rigorous preprocessing of individual omics datasets. This includes:
- Normalization to account for technical variation
- Missing value imputation using appropriate algorithms
- Batch effect correction to remove non-biological variation
- Data scaling to make different omics measurements comparable
Quality assessment should include both technical replicates and internal standards to ensure data reliability across all analytical platforms.
Pathway Enrichment Analysis Protocol
The following step-by-step protocol describes integrated pathway enrichment analysis:
Step 1: Data Preparation
- Generate normalized omics datasets for each molecular domain
- Create ranked lists of molecules based on differential expression or concentration across cycle phases
- Ensure consistent molecule identifiers across all datasets
Step 2: Pathway Enrichment Analysis
- For threshold-based analysis using g:Profiler:
- Input statistically significant molecule lists
- Set functional category size limits (recommended: 5-350 genes/molecules)
- Set query/term intersection threshold (recommended: ≥3 molecules)
- Select relevant pathway databases (GO Biological Processes, Reactome) [44]
- For rank-based analysis using GSEA:
- Input complete ranked lists of all measured molecules
- Select appropriate permutation type (recommended: phenotype for menstrual cycle studies)
- Set significance thresholds (FDR < 0.25, NOM p-value < 0.05) [45]
Step 3: Results Interpretation
- Filter significantly enriched pathways using multiple testing correction (FDR < 0.05)
- Identify pathways with coordinated changes across multiple omics layers
- Interpret biological themes in context of menstrual cycle physiology
Figure 1: Multi-Omics Pathway Analysis Workflow
Visualization and Interpretation
EnrichmentMap Visualization
EnrichmentMap is a Cytoscape app that creates similarity networks of enriched pathways, effectively reducing redundancy by grouping related gene sets [46]. The visualization process involves:
- Loading enrichment results from g:Profiler or GSEA into Cytoscape
- Building the similarity network where nodes represent pathways and edges represent gene overlap
Adjusting display parameters to optimize information density:
- Node size proportional to number of molecules in pathway
- Edge thickness proportional to degree of molecular overlap
- Node color indicating phase-specific enrichment [46]
Cluster identification using automated algorithms (e.g., clusterMaker2)
- Cluster annotation with representative labels using AutoAnnotate
This approach effectively addresses the challenge of interpreting large sets of enriched pathways, which commonly occur in menstrual cycle studies due to the pervasive nature of hormonal regulation.
Integrated Pathway Modeling
For menstrual cycle research, integrated pathway modeling should focus on metabolic pathways showing rhythmicity across cycle phases. Key pathways of interest include:
- Glutathione metabolism - Shows significant variation across menstrual phases [1]
- Urea cycle - Demonstrates phase-specific activity patterns [1]
- Phosphatidylinositol signaling - PI(36:2) identified as potential menstrual pain biomarker [5]
- Amino acid metabolism - Multiple amino acids decrease significantly during luteal phase [1]
Figure 2: Key Pathways in Menstrual Cycle Rhythmicity
Application to Amino Acid-Lipid Rhythmicity
Metabolic Rhythmicity Across the Menstrual Cycle
Comprehensive metabolic profiling has revealed significant oscillations in amino acid and lipid species across the menstrual cycle. A study of 34 healthy women found 39 amino acids and derivatives and 18 lipid species that significantly decreased during the luteal phase (FDR < 0.20), potentially indicating an anabolic state during the progesterone peak with recovery during menstruation and follicular phases [1].
Table 2: Significant Metabolic Changes Across Menstrual Cycle Phases
| Metabolite Class | Change Pattern | Specific Examples | Proposed Biological Significance |
|---|---|---|---|
| Amino Acids | Decrease in luteal phase | Ornithine, arginine, alanine, glycine, methionine, proline | Increased protein synthesis or nitrogen utilization during progesterone peak |
| Phospholipids | Decrease in luteal phase | LPCs, PCs, LPEs | Membrane remodeling or signaling precursor utilization |
| BCAAs | Inverse correlation with pain | Valine, leucine, isoleucine | Decreased levels in severe menstrual pain |
| Phosphatidylinositols | Positive correlation with pain | PI(36:2), PI(38:3), PI(38:4) | Increased levels in severe menstrual pain; potential signaling role |
| Vitamin D | Increase in menstrual phase | 25-OH vitamin D | Phase-specific utilization or regulation |
The rhythmicity of these metabolic pathways may represent periods of increased vulnerability to hormone-related health issues such as premenstrual syndrome (PMS) and premenstrual dysphoric disorder (PMDD) [1].
Biomarker Discovery for Menstrual Symptoms
Integrated multi-omics approaches have identified novel biomarker ratios that effectively discriminate between menstrual pain severity levels. Specifically, the ratio of phosphatidylinositol PI(36:2) to branched-chain amino acids shows remarkable diagnostic accuracy:
- PI(36:2)/BCAA ratio: AUC = 0.990 for discriminating severe vs. mild pain
- PI(36:2)/leucine ratio: AUC = 0.979
- PI(36:2)/valine ratio: AUC = 0.969
- PI(36:2)/isoleucine ratio: AUC = 0.969 [5]
These biomarker ratios demonstrate consistent discrimination accuracy across all menstrual cycle phases, suggesting potential utility for both prediction and monitoring of menstrual pain severity.
Research Reagent Solutions
Table 3: Essential Research Reagents for Multi-Omics Menstrual Cycle Studies
| Reagent Category | Specific Products/Platforms | Application | Key Features |
|---|---|---|---|
| Pathway Analysis Tools | g:Profiler, GSEA, EnrichmentMap | Statistical enrichment analysis | Multiple testing correction, visualization capabilities |
| Metabolomics Platforms | LC-MS, GC-MS systems | Small molecule quantification | Broad metabolite coverage, high sensitivity |
| Lipidomics Reagents | Lipid extraction kits, internal standards | Lipid species identification and quantification | Comprehensive coverage of lipid classes |
| Hormone Assays | ELISA kits, LC-MS/MS assays | Estradiol, progesterone, LH, FSH measurement | Precise cycle phase determination |
| Pathway Databases | GO, Reactome, MSigDB | Biological context interpretation | Curated pathway information |
| Biofluid Collection | Finger-prick blood collection devices | Minimally invasive sampling | Enables frequent at-home sampling |
Integrated multi-omics strategies for pathway analysis provide powerful methodological frameworks for elucidating the complex metabolic rhythmicity of the menstrual cycle. By combining data from multiple molecular domains, researchers can identify coordinated pathway fluctuations that would remain invisible in single-omics approaches. The application of these methods has already revealed significant insights into amino acid and lipid rhythmicity, identifying potential biomarkers for menstrual pain severity and revealing systematic metabolic changes across cycle phases.
As these methodologies continue to evolve, they hold promise for developing personalized interventions for menstrual-related disorders and advancing our understanding of female physiology. The integration of additional omics layers, including proteomics and genomics, with advanced computational modeling will further enhance our ability to decode the complex interactions between hormonal regulation and metabolic pathways throughout the menstrual cycle.
The discovery of robust biomarkers is pivotal for advancing diagnostic and therapeutic strategies in modern medicine. This whitepaper delineates the sophisticated journey from identifying broad metabolic patterns to deriving precise diagnostic ratios, with a specific focus on the dynamic interplay between amino acids and lipids throughout the menstrual cycle. For researchers and drug development professionals, understanding this rhythmicity is not merely an academic exercise but a fundamental requirement for developing accurate diagnostic tools and effective, phase-specific therapeutics for women's health. The cyclical variation in these metabolites, once considered noise, is now emerging as a critical source of biological signal, offering a novel paradigm for biomarker discovery and application.
Biological rhythms are ubiquitous in human physiology, governing processes from cellular metabolism to systemic hormonal release. The menstrual cycle represents a quintessential biological rhythm, driven by the intricate interplay of progesterone, estradiol, follicular stimulating hormone (FSH), and luteinizing hormone (LH) [1] [8]. Beyond its reproductive function, this cycle exerts a profound influence on the body's metabolic landscape. Recent advances in metabolomics have begun to systematically characterize these changes, revealing that the menstrual cycle is a key variable in the metabolic phenotype of premenopausal women [1]. Acknowledging and quantifying these rhythmic variations is the first crucial step in transforming them from confounding factors into powerful diagnostic and therapeutic assets.
Foundational Metabolic Patterns in the Menstrual Cycle
Comprehensive metabolic profiling has unveiled that a significant proportion of the metabolome oscillates during a healthy menstrual cycle. In one pivotal study, 208 out of 397 tested metabolites and micronutrients showed significant changes (p < 0.05), with 71 meeting a False Discovery Rate (FDR) threshold of q < 0.20, underscoring the pervasiveness of this metabolic rhythmicity [1] [8].
Table 1: Summary of Key Metabolic Changes Across the Menstrual Cycle
| Metabolite Class | Specific Metabolites | Direction of Change | Phase of Most Significant Change |
|---|---|---|---|
| Amino Acids & Biogenic Amines | Ornithine, Arginine, Alanine, Glycine, Methionine, Proline, Threonine, Serine | Decrease | Luteal Phase [1] [8] |
| Phospholipids | LPCs, PCs, LPE 22:6 | Decrease | Luteal Phase [1] |
| Vitamins & Cofactors | 25-OH Vitamin D, Pyridoxic Acid | Increase | Menstrual Phase [1] |
| Acylcarnitines | Multiple species | Increase | Periovulatory Phase [1] |
Amino Acid Rhythmicity
The most pronounced cyclic pattern is observed in the plasma amino acid pool. Targeted analyses have consistently found that levels of numerous amino acids and their derivatives decline during the luteal phase. In a detailed study of 34 healthy women, 39 amino acids and derivatives were significantly decreased (FDR < 0.20) in the luteal phase compared to the menstrual and follicular phases [1] [8]. Key amino acids like ornithine, arginine, alanine, and glycine showed statistical significance across multiple phase contrasts (luteal-follicular, luteal-menstrual, luteal-periovulatory) [8]. This collective reduction may indicate a state of increased protein synthesis or heightened nitrogen utilization during the post-ovulatory period, potentially in preparation for a potential pregnancy [1].
Lipid Rhythmicity
Concurrent with amino acid fluctuations, specific lipid species also demonstrate menstrual cycle dependency. Lipidomic analyses reveal that numerous phospholipids, including lysophosphatidylcholines (LPCs), phosphatidylcholines (PCs), and lysophosphatidylethanolamines (LPEs), follow a pattern similar to amino acids, with concentrations dropping in the luteal phase [1]. Of the 139 lipid species tested, 57 were significantly altered, with 17 meeting the FDR threshold for the luteal-follicular contrast [1]. This coordinated decrease in amino acids and lipids suggests a systemic, hormone-driven shift in anabolic metabolism.
Diagram 1: Metabolic Shift in the Luteal Phase. This diagram illustrates the proposed cascade from hormonal changes to observed metabolic patterns.
From Patterns to Diagnostic Ratios: A Case Study in Menstrual Pain
The identification of broad metabolic patterns is foundational, but the true power of biomarker discovery lies in distilling these complex patterns into simple, robust, and clinically actionable diagnostic ratios. A recent 2025 study on menstrual pain (dysmenorrhea) provides a seminal example of this evolution [5].
This research employed a multi-platform metabolomics approach on finger-prick blood samples to identify biomarkers that could objectively differentiate between mild and severe menstrual pain. The initial profiling stage identified several candidate metabolites, including the phosphatidylinositol PI(36:2), which was elevated in the severe pain group, and the branched-chain amino acids (BCAAs) valine, leucine, and isoleucine, which were lower [5]. While each of these metabolites individually showed good discriminatory power in a Receiver Operating Characteristic (ROC) analysis, their diagnostic performance was significantly enhanced when combined into a ratio.
Table 2: Performance of Individual Metabolites vs. Ratios for Discriminating Menstrual Pain Severity
| Biomarker | Area Under Curve (AUC) | Fold Change in Severe vs. Mild Pain |
|---|---|---|
| PI(36:2) | 0.906 | 1.42 increase [5] |
| Valine | 0.865 | ~0.8 decrease [5] |
| Leucine | 0.865 | ~0.8 decrease [5] |
| Isoleucine | 0.865 | ~0.8 decrease [5] |
| PI(36:2)/Total BCAA | 0.990 | N/A [5] |
| PI(36:2)/Leucine | 0.979 | N/A [5] |
The ratio of PI(36:2) to total BCAAs achieved a near-perfect AUC of 0.990, demonstrating exceptional accuracy in classifying pain severity. Furthermore, this ratio showed a significant positive linear correlation with the Numerical Pain Rating Scale (NPRS) and maintained high discrimination accuracy across all phases of the menstrual cycle, making it a reliable predictive and monitoring tool [5].
Experimental Protocols for Metabolic Biomarker Discovery
Reproducible discovery of metabolic patterns and ratios requires rigorous, standardized protocols. The following section outlines key methodologies derived from the cited studies.
Sample Collection and Preparation
- Cohort Design: Recruit premenopausal women with confirmed regular, ovulatory cycles. Phase classification should be confirmed via serum hormone measurements (progesterone, estradiol, FSH, LH) and/or urinary luteinizing hormone tests, in addition to self-reported cycle timing [1] [8].
- Sampling Protocol: Collect biofluids (plasma, serum, urine) at multiple, well-defined timepoints across the cycle (e.g., menstrual, follicular, periovulatory, luteal, pre-menstrual phases) [1]. The use of finger-prick blood (∼100 µL) can minimize participant burden and facilitate frequent sampling [5].
- Sample Processing: For comprehensive analysis, split samples for different analytical platforms. Immediately freeze samples at -80°C after processing to preserve labile metabolites [5].
Analytical Techniques for Metabolomics
A multi-platform approach is essential to cover the diverse chemical and concentration ranges of metabolites.
- Liquid Chromatography-Mass Spectrometry (LC-MS/MS): The workhorse for metabolomics and lipidomics. It is highly sensitive and selective for quantifying amino acids, lipids, and other metabolites in complex biological samples [1] [47].
- For Hydrophilic Metabolites (e.g., Amino Acids): Use Hydrophilic Interaction Liquid Chromatography (HILIC) to overcome the poor retention of polar compounds on standard reverse-phase columns [47].
- Gas Chromatography-Mass Spectrometry (GC-MS): Often used for profiling organic acids and other volatile or derivatized metabolites [1].
- Nuclear Magnetic Resonance (NMR) Spectroscopy: Useful for broad-spectrum metabolite profiling and structural elucidation, though generally less sensitive than MS-based methods [47].
Diagram 2: Multi-Platform Metabolomics Workflow. This diagram outlines the parallel analytical paths used for comprehensive metabolite coverage.
Data Processing and Statistical Analysis
- Pre-processing: Perform peak picking, alignment, and normalization on raw data to generate a quantifiable metabolite data matrix.
- Statistical Analysis:
- Univariate Analysis: Use tests like ANOVA or t-tests to identify metabolites with significant concentration changes between phases or groups (p-value < 0.05) [1] [8].
- Multiple Testing Correction: Apply methods like Benjamini-Hochberg to control the False Discovery Rate (FDR) and minimize false positives (e.g., q < 0.20) [1].
- Multivariate Analysis: Employ Principal Component Analysis (PCA) and Partial Least Squares-Discriminant Analysis (PLS-DA) to visualize group separations and identify metabolite patterns [5].
- Diagnostic Performance: Use Receiver Operating Characteristic (ROC) curve analysis to evaluate the ability of individual metabolites or their ratios to classify clinical outcomes [5].
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Key Reagent Solutions for Menstrual Cycle Metabolomics Research
| Reagent / Material | Function / Application | Technical Notes |
|---|---|---|
| LC-MS/MS Grade Solvents | Mobile phase for chromatography; sample extraction. | High purity is critical to minimize background noise and ion suppression. |
| Stable Isotope-Labeled Internal Standards | Normalization for MS quantification; correct for recovery and matrix effects. | Essential for precise absolute quantification of amino acids and lipids [47]. |
| HILIC Chromatography Columns | Separation of highly polar metabolites like underivatized amino acids. | Preferred over derivatization for its simplicity and compatibility with MS [47]. |
| Derivatization Reagents | Chemically modify metabolites to enhance MS detection sensitivity or enable chiral separation. | e.g., DMT-(S)-Pro-OSu for separating D/L-amino acid enantiomers [47]. |
| Solid Phase Extraction (SPE) Kits | Fractionate and purify complex biological samples prior to analysis. | Can separate lipid classes or remove interfering salts and proteins. |
| Certified Reference Material | Calibrate instruments and validate analytical methods. | Standardized mixtures of known metabolite concentrations. |
The journey from mapping metabolic patterns to validating diagnostic ratios represents a maturation of biomarker science. Research into amino acid and lipid rhythmicity throughout the menstrual cycle has moved beyond cataloging changes and is now yielding specific, high-performance ratios with direct clinical relevance, as demonstrated by the PI(36:2)/BCAA index for menstrual pain. For the drug development community, these findings are transformative. They underscore the necessity of stratifying clinical trial populations and pharmacokinetic analyses by menstrual cycle phase to reduce data variability and uncover true therapeutic efficacy. Future work will focus on validating these ratios in larger, diverse populations and integrating them with other omics data layers to build a systems-level understanding of female physiology, ultimately paving the way for more personalized and effective healthcare for women.
Navigating Research Complexities: Troubleshooting in Menstrual Cycle Metabolomics
The menstrual cycle represents a complex, rhythmic physiological process governed by interacting levels of progesterone, estradiol, follicular stimulating, and luteinizing hormones. Recent advances in metabolomics have revealed significant cyclic fluctuations in plasma amino acids, lipids, and other metabolites throughout its phases [1]. Of 397 metabolites tested in one comprehensive study, 208 showed significant changes (p < 0.05) across the menstrual cycle, with 71 meeting the false discovery rate threshold of 0.20 [1]. These fluctuations create substantial inter-individual variability in symptom experiences and metabolic responses that complicate research and therapeutic development.
This technical guide addresses these challenges by providing a framework for stratification based on symptom burden and skin type, enabling researchers to account for variability in study design and analysis. By implementing these approaches, scientists can advance personalized interventions in women's health, particularly in the context of amino acid and lipid rhythmicity research.
Metabolic Rhythmicity of Amino Acids and Lipids Across the Menstrual Cycle
Phase-Dependent Fluctuations in Metabolic Profiles
Comprehensive metabolomic profiling has established that the menstrual cycle exhibits predictable rhythmicity in amino acid and lipid metabolism. A study of 34 healthy premenopausal women collected biofluids at four timepoints and classified cycles into five phases: menstrual (M), follicular (F), periovular (O), luteal (L), and pre-menstrual (P) [1]. The analysis revealed consistent patterns of metabolite changes across these phases.
Table 1: Significant Metabolite Changes Across Menstrual Cycle Phases [1]
| Metabolite Class | Total Compounds | Significantly Changed (p<0.05) | Key Findings | Primary Phase Associations |
|---|---|---|---|---|
| Amino Acids & Derivatives | 54 | 48 | 37 amines significant in L-M contrast (q<0.20) | Decreased in luteal phase |
| Phospholipids | 139 | 57 | 17 lipid species met FDR threshold for L-F | Decreased in luteal phase |
| Clinical Parameters & Vitamins | 27 | 6 | Vitamin D elevated in menstrual phase | Increased in menstrual phase |
| Acylcarnitines | 50 | 19 (plasma), 16 (urine) | Majority increased in periovulatory phase | Increased in periovulatory phase |
The data reveals a consistent pattern of metabolic suppression during the luteal phase, with 39 amino acids and derivatives and 18 lipid species demonstrating significant decreases (FDR < 0.20) [1]. This pattern may represent an anabolic state during the progesterone peak, with recovery occurring during menstruation and the follicular phase. These rhythmic fluctuations establish the foundation for understanding inter-individual variability in symptom experiences.
Symptom-Associated Metabolic Biomarkers
Beyond phase-specific changes, recent research has identified specific metabolite profiles associated with menstrual symptom severity. A 2025 study investigating menstrual pain identified branched-chain amino acids (BCAAs) and specific phosphatidylinositol (PI) species as biomarkers differentiating pain severity [5].
Table 2: Metabolite Biomarkers of Menstrual Pain Severity [5]
| Metabolite | Change in Severe vs Mild Pain | AUC Value | Biological Significance |
|---|---|---|---|
| PI(36:2) | 1.42-fold increase | 0.906 | Cell signaling membrane composition |
| Valine | 0.8-fold decrease | 0.865 | Branched-chain amino acid metabolism |
| Isoleucine | 0.8-fold decrease | 0.865 | Branched-chain amino acid metabolism |
| Leucine | 0.8-fold decrease | 0.865 | Branched-chain amino acid metabolism |
| PI(36:2)/BCAA ratio | Inverse correlation | 0.990 | Enhanced discrimination accuracy |
The ratio of PI(36:2) to BCAAs demonstrated remarkable discriminatory power, with area under the curve (AUC) values reaching 0.990 during the menstrual phase [5]. These ratios maintained high discrimination accuracy across all menstrual cycle phases and showed significant positive linear correlation with numerical pain rating scale (NPRS) scores, supporting their use for objective pain assessment in research and clinical applications [5].
Stratification Framework: Accounting for Inter-individual Variability
Symptom Burden Stratification
The symptom burden represents a more significant determinant of functional outcomes than menstrual phase alone. Research across multiple populations demonstrates that symptom frequency and severity better predict sleep quality, recovery-stress states, and performance metrics than hormonal phases [19].
In elite female basketball athletes, higher daily symptom burden and greater overall symptom frequency consistently correlated with poorer sleep quality, reduced recovery, and elevated stress, while menstrual cycle phases showed only limited and inconsistent associations [19]. This suggests that stratification should prioritize symptom burden quantification over phase-based grouping alone.
Stratification Protocol:
- Baseline Symptom Assessment: Implement validated instruments (NPRS, MDQ, DRSP) during screening
- Daily Symptom Tracking: Collect daily symptom data across at least one complete menstrual cycle
- Symptom Burden Quantification: Calculate composite scores incorporating frequency, severity, and functional impact
- Stratification Thresholds: Establish population-specific percentiles (e.g., mild: <25th percentile; severe: >75th percentile)
Skin Type Considerations in Metabolic Research
Skin type and permeability represent critical variables in metabolomic studies, particularly those utilizing minimally-invasive sampling methods. Recent methodological advances enable comprehensive metabolic profiling from minimal blood volumes (100μL finger-prick samples) [5], but these approaches may be influenced by skin barrier function and composition.
Key Stratification Variables for Skin Type:
- Skin Barrier Integrity: Measured via transepidermal water loss (TEWL)
- Permeability Characteristics: Particularly relevant for transdermal sampling methodologies
- Inflammatory Status: Assessed through CRP and cytokine profiling
- Demographic Factors: Fitzpatrick skin type, age, and photodamage history
Experimental Protocols for Stratified Research Design
Comprehensive Metabolic Profiling Protocol
The following protocol enables robust metabolic profiling across menstrual cycle phases while accounting for inter-individual variability through stratified sampling:
Participant Recruitment & Screening:
- Include regularly cycling women (25-38 day cycles) aged 18-45
- Exclude hormonal contraceptive use, unstable medical/psychiatric conditions, and recent medication changes
- Document skin type using standardized classification systems
- Conduct baseline symptom burden assessment
Sample Collection & Processing:
- Phase Verification: Confirm cycle phase through serum hormones, urinary luteinizing hormone, and self-reported timing [1]
- Blood Collection: Collect 100μL finger-prick blood at five phase timepoints (menstrual, follicular, periovular, luteal, pre-menstrual) [5]
- Sample Processing: Immediately process samples using validated pretreatment procedures for multiple analytical methods [5]
- Symptom Documentation: Record concurrent symptom burden using standardized instruments
Analytical Methodology:
- Metabolomics: LC-MS and GC-MS for comprehensive metabolite profiling [1]
- Lipidomics: Targeted analysis of phospholipid species, particularly phosphatidylinositol [5]
- Hormone Assays: LC-MS/MS for steroid hormone quantification [5]
- Clinical Chemistries: Standard enzymatic/colorimetric methods for CRP, lipids, vitamins [1]
Data Analysis Framework for Stratified Outcomes
Statistical Approach:
- Implement linear mixed modeling to account for repeated measures and intra-individual variation [19]
- Conduct phase-phase comparisons with correction for multiple testing (FDR < 0.20) [1]
- Employ multivariate analyses (PCA, PLS-DA) to identify global metabolite patterns [5]
- Perform ROC analysis to evaluate biomarker discriminatory power [5]
Stratification Analysis:
- Primary Analysis: Compare metabolic patterns across menstrual cycle phases
- Stratified Analysis: Conduct subgroup analyses based on symptom burden severity
- Interaction Testing: Evaluate phase × symptom burden interaction effects
- Predictive Modeling: Develop algorithms predicting symptom severity from metabolic profiles
Visualization of Metabolic Rhythmicity and Symptom Relationships
Diagram 1: Integrated research workflow for stratified menstrual cycle metabolomics.
Diagram 2: Key metabolic pathways in menstrual symptom expression showing biomarker relationships.
Research Reagent Solutions for Menstrual Cycle Metabolomics
Table 3: Essential Research Reagents for Menstrual Cycle Metabolomics
| Reagent Category | Specific Products | Application | Technical Considerations |
|---|---|---|---|
| Hormone Assay Kits | LC-MS/MS steroid panels, ELISA for LH/FSH | Cycle phase verification | Cross-reactivity assessment, sensitivity <5 pg/mL for estradiol |
| Metabolomics Standards | Cambridge Isotope labeled internal standards | Quantification of 692+ metabolites [5] | Coverage of amino acids, lipids, organic acids |
| Sample Collection | Finger-prick blood collection devices, SALIVette swabs | Minimally-invasive sampling | Compatibility with 100μL volumes [5] |
| Lipidomics Reagents | Phosphatidylinositol standards, lipid extraction kits | Targeted PI species quantification | Focus on PI(36:2), PI(38:3), PI(38:4) [5] |
| Symptom Assessment | Validated questionnaires (NPRS, DLQI) | Symptom burden quantification | Daily tracking capability, validation in target population |
The integration of symptom burden stratification with comprehensive metabolic profiling represents a paradigm shift in menstrual cycle research. By moving beyond phase-based grouping alone to incorporate individualized symptom metrics and skin type considerations, researchers can account for significant inter-individual variability in study outcomes. The biomarker ratios identified in recent research, particularly the PI(36:2)/BCAA index with AUC values reaching 0.990 [5], offer promising tools for objective symptom assessment and personalized intervention development.
Future research directions should focus on validating these stratification approaches across diverse populations, developing standardized protocols for symptom burden quantification, and exploring the molecular mechanisms linking specific metabolite patterns to symptom experiences. Through these advances, the field can progress toward truly personalized interventions that address the unique physiological and symptomatic experiences of individual women across their menstrual cycles.
Accurately verifying menstrual cycle phases presents a significant methodological challenge in female-specific research, particularly in studies investigating the rhythmicity of amino acids and lipids. This technical guide examines the gold standard methodologies for phase verification and explores practical, evidence-based compromises for research settings where ideal conditions are not feasible. By synthesizing current literature and emerging technologies, this whitepaper provides researchers and drug development professionals with a structured framework for implementing rigorous menstrual cycle tracking while maintaining scientific validity across different research environments. The critical importance of precise phase verification is highlighted through its essential role in generating reliable data on metabolic fluctuations throughout the menstrual cycle.
The menstrual cycle represents a fundamental biological rhythm characterized by complex interactions between ovarian, hormonal, and endometrial cycles that create a dynamic physiological environment [48]. For researchers investigating amino acid and lipid rhythmicity, these fluctuations are particularly significant as estrogen and progesterone directly regulate various aspects of metabolism, including substrate utilization, energy expenditure, and insulin sensitivity [49]. The challenge for researchers lies in accurately capturing these metabolic changes within their specific hormonal contexts, which requires precise determination of menstrual cycle phases.
Unfortunately, the field faces a significant methodological crisis, with many studies relying on assumed or estimated menstrual cycle phases rather than direct verification [48]. This approach amounts to guessing the occurrence and timing of ovarian hormone fluctuations and risks generating invalid and unreliable data with potentially significant implications for understanding female athlete health, training, performance, and injury [48]. The problem is particularly acute in metabolic studies, where research has demonstrated that amino acids and phospholipids show significant phase-dependent variations that could be overlooked with imprecise phase verification [1] [50].
This whitepaper establishes a comprehensive framework for overcoming phase verification challenges by presenting both gold standard methodologies and practical compromises suitable for various research contexts. By addressing terminology standardization, validation techniques, and implementation strategies specific to amino acid and lipid rhythmicity research, we aim to elevate methodological rigor in female-focused physiological studies.
Gold Standard: Direct Verification Methodologies
Defining the Gold Standard
The gold standard for menstrual cycle phase verification in research contexts involves a multi-modal approach that combines direct physiological measurements with hormonal assays to confirm both the timing of ovulation and the resulting hormonal milieu [51] [48]. This comprehensive methodology transcends traditional calendar-based counting, which fails to detect subtle menstrual disturbances such as anovulatory or luteal phase deficient cycles that present with meaningfully different hormonal profiles [48]. A eumenorrheic cycle suitable for research purposes should be characterized by cycle lengths ≥ 21 days and ≤ 35 days, evidence of a luteinizing hormone surge, and the correct hormonal profile with sufficient progesterone elevation during the luteal phase [48].
The Quantum Menstrual Health Monitoring Study exemplifies this gold standard approach by measuring four key reproductive hormones in urine - follicle-stimulating hormone (FSH), estrone-3-glucuronide (E13G), luteinizing hormone (LH), and pregnanediol glucuronide (PDG) - to characterize patterns that predict and confirm ovulation, referenced to serum hormones and the gold standard of the ultrasound day of ovulation [51]. This integrated protocol establishes a rigorous framework for phase verification that can be specifically adapted for metabolic rhythmicity research.
Experimental Protocols for Gold Standard Verification
Implementing gold standard phase verification requires meticulous experimental design with specific protocols for different menstrual cycle phases:
Ovulation Confirmation Protocol: The Quantum Study protocol involves participants tracking their menstrual cycles for 3 months while using an at-home quantitative urine hormone monitor (Mira monitor) to predict ovulation [51]. The day of ovulation is then confirmed with serial transvaginal ultrasounds completed in a clinical setting, with urine results compared to serum hormone values [51]. This direct ultrasound visualization provides the most definitive confirmation of ovulation occurrence.
Luteal Phase Verification Protocol: To confirm adequate luteal phase function, testing should occur 7-9 days post-LH surge detection to capture peak progesterone levels [49]. This timing ensures the exclusion of anovulatory participants and those with luteal phase defects who would not exhibit appropriate progesterone elevation. Serum progesterone levels provide the most direct assessment, though validated saliva testing or urinary PDG measurements offer acceptable alternatives in certain research contexts.
Multi-Phase Sampling Protocol: For comprehensive metabolic profiling across cycle phases, researchers should establish five distinct sampling points aligned with hormonal status: menstrual phase (days 1-5), late follicular phase (days 7-10), periovulatory phase (LH surge), early luteal phase (days 2-4 post-ovulation), and mid-luteal phase (days 7-9 post-ovulation) [1]. This sampling strategy captures metabolic states across the entire hormonal spectrum.
Table 1: Gold Standard Phase Verification Methods
| Verification Component | Gold Standard Method | Alternative Methods | Validation Requirements |
|---|---|---|---|
| Ovulation Detection | Serial follicular-tracking ultrasound | Urinary LH surge testing | Daily testing until positive |
| Luteal Phase Confirmation | Serum progesterone ≥ 5 ng/mL | Urinary PDG, saliva progesterone | Sampling 7-9 days post-LH surge |
| Cycle Regularity | 3-month bleeding calendar + hormonal confirmation | Cycle length history (21-35 days) | Exclusion of anovulatory cycles |
| Hormonal Status | Serum E2 and P4 measurements | Quantitative urine hormone monitoring | Phase-specific threshold values |
Research Reagent Solutions for Gold Standard Verification
Table 2: Essential Research Reagents and Materials for Phase Verification
| Reagent/Material | Function in Phase Verification | Implementation Example |
|---|---|---|
| Urinary LH Test Kits | Detects LH surge preceding ovulation | Home testing starting cycle day 8 |
| Mira Fertility Monitor & Wands | Quantitative measurement of FSH, E13G, LH, PDG | At-home daily tracking across cycle |
| Serum Progesterone Immunoassays | Confirms ovulatory cycle and luteal function | Clinical lab testing 7-9 days post-LH surge |
| Ultrasound Equipment | Visualizes follicular development and rupture | Clinical setting for ovulation confirmation |
| HPLC-MS/MS Systems | Quantifies steroid hormones in serum/urine | Laboratory hormone validation |
| Salivary Hormone Kits | Non-invasive progesterone measurement | Field-based luteal phase verification |
Practical Compromises: Evidence-Based Alternatives
Tiered Verification Framework
While gold standard methods represent the ideal approach, practical constraints often necessitate evidence-based compromises. A tiered verification framework provides structured alternatives while maintaining scientific rigor:
Tier 1 (Comprehensive Verification): Incorporates urinary hormone monitoring with quantitative devices (e.g., Mira monitor) combined with occasional serum validation and structured symptom tracking [51] [49]. This approach maintains high accuracy while reducing participant burden compared to daily ultrasound monitoring.
Tier 2 (Moderate Verification): Utilizes urinary LH surge detection kits to identify ovulation, combined with basal body temperature tracking to confirm luteal phase shift and calendar-based estimation for phase timing [49]. This approach provides reasonable accuracy for many research contexts without requiring specialized equipment.
Tier 3 (Minimum Verification): Relies on calendar-based counting of cycle days with exclusion of participants reporting irregular cycles, combined with structured menstrual symptom questionnaires [48]. This approach should include transparent reporting of limitations and acknowledge the potential inclusion of anovulatory cycles.
Technological Innovations in Phase Verification
Recent technological advances have created new opportunities for practical yet accurate phase verification:
Quantitative Urinary Hormone Monitors: Devices like the Mira monitor provide quantitative measurements of key reproductive hormones (FSH, E13G, LH, PDG) at home, generating numerical values that can be tracked over time [51]. These systems incorporate algorithmic analysis to identify hormone patterns predictive of ovulation and confirm luteal phase status.
Integrated Digital Platforms: Several systems now combine hormonal data with secondary parameters including basal body temperature, resting heart rate, heart rate variability, and self-reported symptoms to improve phase prediction accuracy [19]. These multimodal approaches can enhance reliability when direct hormonal confirmation is not feasible.
Minimally Invasive Sampling Techniques: Recent methodological advances enable comprehensive metabolic profiling from minimal sample volumes, with one study quantifying 692 metabolites from only 100μL of finger-prick blood [50]. Such approaches facilitate frequent sampling across multiple cycle phases while reducing participant burden.
Table 3: Practical Compromise Approaches for Different Research Settings
| Research Context | Recommended Compromise | Key Methodological Elements | Limitations & Mitigations |
|---|---|---|---|
| Elite Sport Field Studies | Tier 1: Quantitative urine hormone monitoring | Mira monitor + symptom tracking + cycle history | Limited serum validation; report as estimated hormonal status |
| Large Cohort Epidemiological Studies | Tier 2: LH surge detection + calendar tracking | Urinary LH kits + cycle day counting | Some anovulatory cycles included; report percentage with confirmed ovulation |
| Community-Based Exercise Studies | Tier 3: Calendar-based + symptom questionnaires | Cycle regularity screening + Mansfield-Voda-Jorgensen Bleeding Scale | High rate of undetected anovulatory cycles; transparent limitations reporting |
| Metabolic Rhythmicity Lab Studies | Tier 1 with frequent blood sampling | Quantitative hormones + frequent metabolite sampling | Participant burden; consider finger-prick blood collection methods |
Methodological Implementation for Amino Acid and Lipid Rhythmicity
Phase-Specific Metabolic Sampling Protocols
Research investigating amino acid and lipid rhythmicity across the menstrual cycle requires specialized sampling protocols designed to capture phase-specific metabolic fluctuations:
Targeted Sampling Points: Based on metabolic studies showing significant phase-dependent variations, researchers should prioritize sampling during five key phases: (1) early follicular (menstruation), (2) late follicular (pre-ovulatory), (3) periovulatory (LH surge), (4) early luteal, and (5) mid-luteal phases [1]. This approach captures the dynamic metabolic transitions occurring across the cycle.
Frequent Sampling Design: For detailed rhythmicity analysis, studies should incorporate more frequent sampling (every 2-4 days) throughout the cycle, with alignment based on confirmed hormonal events rather than calendar days [32]. This approach enables precise characterization of metabolic trajectories rather than single timepoint comparisons.
Standardized Pre-Analytical Processing: Given the sensitivity of metabolomic measurements, standardized protocols for sample collection, processing, and storage are essential. This includes consistent fasting status, time of day collection, immediate processing, and uniform storage at -80°C [1] [50].
Analytical Considerations for Metabolic Rhythmicity
Mass Spectrometry-Based Metabolomics: Comprehensive metabolic profiling requires multiple analytical platforms to capture the diverse chemical properties of amino acids and lipid species. As demonstrated in recent studies, this should include LC-MS for polar metabolites and lipidomics, GC-MS for organic acids, and targeted LC-MS/MS for specialized lipid mediators and steroid hormones [1] [50].
Rhythmicity Analysis Methods: Identifying significant metabolic fluctuations requires appropriate analytical approaches including cosinor analysis with 24-hour periods, linear mixed models to account for within-participant variation, and false discovery rate correction for multiple comparisons [1] [52]. These methods should be pre-specified in analytical plans.
Integration with Hormonal Data: Metabolic measurements must be integrated with hormonal verification data to establish true phase-dependent relationships. Advanced statistical approaches including multivariate analysis and machine learning algorithms can identify complex interactions between hormonal status and metabolic profiles [32] [50].
Research Methodology Workflow: This diagram illustrates the integrated approach necessary for rigorous investigation of amino acid and lipid rhythmicity across verified menstrual cycle phases.
Accurate menstrual cycle phase verification represents a fundamental methodological requirement for rigorous investigation of amino acid and lipid rhythmicity in female participants. While gold standard approaches incorporating direct hormonal measurement and ultrasound confirmation provide the most reliable phase determination, evidence-based compromises can be implemented when practical constraints prevent ideal protocol execution. The critical factor remains transparent reporting of verification methodologies and acknowledgment of limitations when direct measurement is not feasible.
Future methodological development should focus on creating standardized verification protocols specific to metabolic research, validating novel technologies for field-based hormone monitoring, and establishing consensus guidelines for terminology and phase definitions across the research community. By advancing these methodological foundations, researchers can generate more reliable and reproducible data on the complex interactions between menstrual cycle physiology and metabolic rhythmicity, ultimately strengthening the evidence base for female-specific health interventions and pharmaceutical development.
Biological Rhythms Integration: This diagram illustrates the theoretical relationship between central circadian regulation, menstrual cycle hormonal rhythms, and peripheral metabolic outputs that underlie amino acid and lipid rhythmicity.
In the investigation of amino acid and lipid rhythmicity across the menstrual cycle, a profound challenge is the isolation of these dynamics from other powerful, overlapping biological rhythms. Diet, exercise, and endogenous circadian cycles are not merely background variables; they are active, interacting forces that can directly modulate metabolic pathways. Failure to adequately account for these factors can confound results, leading to erroneous conclusions about the temporal patterns of amino acids and lipids. This guide provides researchers and drug development professionals with a technical framework for identifying, controlling, and statistically adjusting for these confounders to ensure the integrity of findings in menstrual cycle research.
Deciphering Key Confounding Factors
The principal confounding factors in this research domain interact in a complex network, influencing the very metabolic processes under investigation. The table below summarizes their core mechanisms and documented impacts on metabolism.
Table 1: Key Confounding Factors and Their Metabolic Influence
| Confounding Factor | Core Mechanism of Action | Documented Impact on Metabolic Rhythms |
|---|---|---|
| Meal Timing [53] [54] | Synchronizes peripheral circadian clocks in organs like the liver, independent of the central brain clock. | Later mealtimes (e.g., first meal after 9 AM, last meal after 9 PM) are associated with adverse cardiovascular outcomes and disrupted glucose metabolism [54]. |
| Dietary Composition [55] [56] | Alters substrate availability for energy production and signaling, and modulates gut-brain axis communication. | "Western" dietary patterns (high in fat, animal protein) are linked to circadian syndrome (CircS) and poorer metabolic health, while "Prudent" patterns (high in carbohydrates, plant-based) are protective [55] [56]. |
| Exercise Timing [57] | Acts as a non-photic zeitgeber, capable of phase-advancing or phase-delaying the internal circadian rhythm. | Morning exercise (10 hours after DLMO) induced a phase advance of 0.62 hours, while evening exercise had a negligible effect. This effect was modified by chronotype [57]. |
| Chronotype [57] [55] | Determines an individual's innate phase relationship between their circadian rhythm and the external light-dark cycle. | Evening chronotypes exhibit distinct dietary patterns, later meal times, and less favorable metabolic profiles (e.g., higher BMI, triglycerides) compared to morning types [55]. |
Experimental Protocols for Control and Measurement
Implementing rigorous methodologies is paramount for isolating the effect of the menstrual cycle from these confounders. The following protocols, drawn from recent studies, provide a template for robust study design.
Protocol for Menstrual Cycle Phase Stratification
A study on sebum composition provides a model for precise phase determination [32].
- Participant Selection: Recruit women (e.g., 24-29 years old) with self-declared cycle durations of 26-30 days. Exclude smokers and those with acne, scarring, or those taking medications that could influence lipid metabolism [32].
- Phase Determination:
- Ovulation Phase (OP): Monitor via commercial ovulation tests starting from day 8 until a positive result is observed.
- Luteal & Follicular Phases: Calculate based on the onset of menstruation and the positive ovulation test. The early luteal phase (ELP) can be defined as the days immediately following ovulation, and the late luteal phase (LLP) as the days preceding the next menstruation [32].
- Sebum/Lipid Sampling: After washing the face with a standardized cleanser and confirming a clean baseline with a Sebumeter (readings <10 μg/cm²), apply Sebutape patches to the forehead and cheeks for 30 minutes. Extract lipids using ethanol with 0.025% BHT (to prevent oxidation) and analyze lipid classes via GC-MS and LC-MS [32].
Protocol for Circadian and Chronotype Assessment
- Chronotype Determination: Utilize the Morningness-Eveningness Questionnaire (MEQ) to categorize participants as morning, intermediate, or evening types [55].
- Objective Circadian Phase Marker: Measure the Dim Light Melatonin Onset (DLMO). This involves collecting saliva samples in dim light (<10-30 lux) every 30-60 minutes in the evening until melatonin levels consistently exceed a threshold (e.g., 4 pg/mL). This provides a precise metric for internal circadian time, which can be used to schedule interventions like exercise [57].
Protocol for Dietary Control and Monitoring
- Dietary Intake Recording: Use weighed food diaries over multiple days (e.g., 7 days) for highly accurate data, as used in chronotype studies [55]. Alternatively, multiple 24-hour dietary records can establish patterns of meal timing [54].
- Key Dietary Variables to Quantify:
The Scientist's Toolkit: Essential Research Reagents
The following table details key materials and reagents essential for executing the methodologies described in this field.
Table 2: Research Reagent Solutions for Metabolic & Circadian Rhythmicity Studies
| Item Name | Function / Application | Technical Notes |
|---|---|---|
| Sebutape Patches (Cuderm S100) | Non-invasive collection of sebum from skin surface for lipidomic analysis [32]. | Patches are applied to forehead/cheeks for 30 min, then stored at -80°C prior to lipid extraction [32]. |
| LC-MS & GC-MS Systems | High-sensitivity quantification of a wide range of metabolites, including lipids (e.g., PIs), amino acids (e.g., BCAAs), and hormones [32] [50]. | GC-MS is ideal for derivatized compounds like FAs and squalene. LC-MS is suited for a broader lipidomics and hydrophilic metabolomics profile [32] [50]. |
| Salivary Melatonin ELISA/Kits | Determination of DLMO for objective circadian phase assessment [57]. | Requires collection under dim light conditions. A threshold of 4 pg/mL is commonly used to define DLMO [57]. |
| Morningness-Eveningness Questionnaire (MEQ) | A standardized survey to classify an individual's innate chronotype [55]. | A higher score indicates a morning chronotype, which is associated with a healthier metabolic profile [55]. |
| Internal Standard Mixture (for MS) | Enables precise quantification of metabolites by accounting for sample preparation variability and instrument drift [32]. | Should include deuterated standards relevant to the analytes of interest (e.g., d6-Cholesterol, d6-Squalene, d5-TG 48:0) [32]. |
Data Synthesis and Statistical Adjustment
When complete control of confounders is not feasible, statistical methods must be employed to account for their effects.
- Regression Modeling: Include confounders like chronotype (MEQ score), meal timing, and physical activity level as covariates in linear or logistic regression models when testing associations between menstrual phase and metabolic outcomes (e.g., amino acid or lipid levels) [55].
- Stratification: Analyze data within subgroups, such as performing separate analyses for morning-type and evening-type participants, to reveal effect modifications that might be masked in the whole cohort [57].
- Interaction Terms: Statistically test for interactions by including product terms (e.g., Menstrual Phase × Chronotype) in regression models to determine if the effect of the menstrual cycle differs based on another factor.
The following diagram illustrates the complex interplay between the core factors discussed and their potential to confound the relationship of primary interest in menstrual cycle research.
Confounding Network in Metabolic Cycle Research: This diagram shows how key confounding factors (red) can independently influence peripheral clocks (blue), which in turn affect the metabolic outcomes of interest. This creates an indirect, non-causal pathway between the menstrual cycle and amino acid/lipid levels, potentially confounding the core relationship (green).
Critical Data Synthesis and Reference Tables
Integrating quantitative findings from disparate studies is crucial for forming hypotheses and powering new experiments.
Table 3: Quantitative Summary of Key Confounding Effects from Recent Literature
| Study Focus | Population | Exposure/Group | Key Quantitative Finding | Effect Size (vs. Reference) |
|---|---|---|---|---|
| Meal Timing & CVD Risk [54] | 103,389 adults (NutriNet-Santé) | Each hour delay of first meal | 6% increased risk of overall cardiovascular disease | HR = 1.06 (1.01–1.12) |
| Last meal after 9 PM (vs. before 8 PM) | 28% increased risk of cerebrovascular disease | HR = 1.28 (1.05–1.55) | ||
| Chronotype & Metabolic Health [55] | 110 Ukrainian adults | 10-point increase in MEQ (more morning-type) | 55% lower risk of being metabolically unhealthy | OR = 0.45 (0.26–0.80) |
| Exercise & Phase Shift [57] | 52 sedentary young adults | Morning Exercise (vs. Evening) | Significant phase advance of circadian rhythm | 0.62 ± 0.18 hours |
| Menstrual Pain Biomarkers [50] | 20 Japanese women | Severe vs. Mild Pain (Menstrual Phase) | PI(36:2) increased, BCAAs decreased | Fold Change: 1.42 (PI), ~0.80 (BCAAs) |
| Ratio PI(36:2)/Total BCAA | Discriminated pain severity with high accuracy | AUC = 0.990 |
In menstrual cycle research focusing on amino acid and lipid rhythmicity, achieving high data quality is paramount. High-throughput technologies, while powerful, magnify the impact of technical noise, which can obscure genuine biological signals, particularly for low-abundance metabolites [58]. This technical noise, stemming from stochastic variation during sequencing, limits of sequencing depth, and alignment inaccuracies, disproportionately affects the quantification of low-abundance molecules [58]. In the context of menstrual cycle studies, where subtle, rhythmic fluctuations in branched-chain amino acids (BCAAs) and specific lipids like phosphatidylinositols are of interest, this noise can completely obscure critical biological patterns [50]. Consistent reduction of this random background noise is therefore not merely a data processing step, but a fundamental prerequisite for capturing functionally meaningful biological signals related to the menstrual cycle.
Understanding and Quantifying Technical Noise
Technical noise in sequencing data is characterized by a lack of consistency and reproducibility. Genes or metabolites whose quantification shows this lack of uniformity are defined as "noisy" [58]. This noise manifests as low-level expression variations that can obscure patterns in downstream analyses. The challenge is particularly acute for low-abundance molecules, which exhibit greater coverage bias and higher susceptibility to technical variability introduced during library preparation and sequencing [58].
In untargeted metabolomics, the problem is magnified by the sheer volume of data. Modern liquid chromatography-mass spectrometry (LC-MS) methods can detect over 25,000 features per sample, but a large proportion of these are non-reproducible artifacts, adducts, and contaminants. Ultimately, this noisy data may yield fewer than 1,000 true metabolites [59]. This noise presents several critical challenges for researchers:
- Subjective Expert Curation: Without standardized methods, different experts may inconsistently classify ambiguous peak groups as real signal or noise, leading to irreproducible results [59].
- Tedious Manual Curation: Manual filtering strategies are time-consuming. Strict thresholds risk losing biologically relevant low-abundant signals, while relaxed thresholds exponentially increase manual curation workload [59].
- Computational Intensity: High-resolution instruments generate gigabytes of data, creating dependencies on high-end computational resources for processing and analysis [59].
The Impact of Noise on Menstrual Cycle Research
The presence of technical noise has direct implications for the study of amino acid and lipid rhythmicity across the menstrual cycle. For instance, a 2025 study investigating metabolic markers for menstrual pain severity identified branched-chain amino acids (BCAAs) and specific phosphatidylinositols (PIs), particularly PI(36:2), as key biomarkers [50]. The study found an inverse correlation where PI(36:2) increased by 1.42-fold in individuals with severe pain, while BCAAs (leucine, isoleucine, and valine) decreased to approximately 0.8-fold [50]. These subtle but biologically critical differences could easily be lost amidst technical noise without proper filtering protocols.
Table 1: Key Metabolite Changes Associated with Menstrual Pain Severity
| Metabolite Class | Specific Metabolite | Change in Severe Pain Group (Fold Change) | Biological Significance |
|---|---|---|---|
| Phosphatidylinositol | PI(36:2) | +1.42 | Potential indicator of inflammatory processes or cellular signaling changes |
| Branched-Chain Amino Acid | Leucine | ~0.8 | May reflect altered energy metabolism or muscle protein turnover |
| Branched-Chain Amino Acid | Isoleucine | ~0.8 | Similar role to leucine in energy and metabolism |
| Branched-Chain Amino Acid | Valine | ~0.8 | Completes the trio of BCAAs showing consistent reduction |
Methodologies for Noise Reduction and Signal Enhancement
ThenoisyRNoise-Filtering Pipeline
The noisyR package provides a comprehensive, data-driven solution for assessing variation in signal distribution and achieving optimal information-consistency across replicates and samples [58]. This selection process facilitates meaningful pattern recognition outside the background-noise range. The package is applicable to both count matrices and sequencing data, outputting sample-specific signal/noise thresholds and filtered expression matrices [58].
The core methodology of noisyR involves quantifying noise based on the correlation of expression across subsets of genes in different samples and replicates, evaluated across all gene abundances [58]. This approach is robust and does not rely on specific biological assumptions, making it suitable for large-scale sequencing efforts where the low number of replicates hinders imputation-based approaches [58].
Machine Learning-Based Solutions for Metabolomics
Machine learning offers a powerful alternative to manual curation for distinguishing true signals from noise in untargeted metabolomics data. One advanced solution, Polly-PeakML, employs a machine learning model trained on thousands of expert-curated peak-groups to automatically classify features as real signal or noise with 94% accuracy [59]. This approach provides consistent, reproducible results and operates approximately 120 times faster than manual curation [59].
This method circumvents the need for optimizing peak-detection parameters, allowing researchers to include low-abundance features without the burden of manually removing noise. This is particularly valuable for capturing the full dynamic range of metabolite concentrations throughout the menstrual cycle.
Experimental Design for Minimizing Noise
Proper experimental design is crucial for minimizing technical noise at its source:
- Sample Collection Consistency: In menstrual cycle research, consistent timing of sample collection relative to cycle phase (e.g., follicular, ovulatory, luteal) is critical [50] [19].
- Minimized Sample Volume: Innovative methods enabling comprehensive profiling from minimal sample volume (e.g., 100 µL of finger-prick blood) reduce participant burden and facilitate longitudinal sampling across multiple cycle phases [50].
- Quality Control Checks: Initial quality checks using tools like FastQC and MultiQC should be performed, followed by density plots, MA plots, and PCA plots to evaluate distribution similarity and identify outliers [58].
Table 2: Essential Reagent Solutions for Low-Abundance Metabolomics
| Research Reagent / Material | Function in Experimental Protocol |
|---|---|
| LC-MS Grade Solvents (Methanol, Isopropanol) | Ensure minimal background interference during metabolite extraction and separation in liquid chromatography. |
| Deuterated Internal Standards (e.g., d6-Cholesterol, d5-TG 48:0) | Correct for technical variability during sample preparation and instrument analysis; enable quantitative accuracy. |
| BSTFA + 1% TCMS (Derivatization Reagent) | Enhances detection of certain metabolite classes (e.g., fatty acids, organic acids) by gas chromatography-mass spectrometry (GC-MS). |
| Ava Fertility Tracker / Salivary Hormone Kits | Objectively verify menstrual cycle phase alongside self-reporting for accurate phase-specific metabolic analysis [19]. |
| Sebutape Patches or Similar | Non-invasive collection of specific biological samples (e.g., sebum) for lipidomic analysis across menstrual cycles [32]. |
| Quality Control (QC) Pool Samples | Pooled aliquots from all study samples used to monitor instrument stability and performance throughout the analytical batch. |
Experimental Protocols for Menstrual Cycle Metabolomics
Comprehensive Metabolomic Profiling Protocol
A robust protocol for studying amino acid and lipid rhythmicity across the menstrual cycle should incorporate multiple analytical methods to capture a wide range of metabolites [50].
Sample Preparation:
- Collection: Collect finger-prick blood (approximately 100 µL) at multiple time points across confirmed menstrual cycle phases (follicular, ovulatory, luteal) [50].
- Extraction: Extract metabolites using ethanol containing 0.025% butylhydroxytoluene (BHT) to prevent oxidation [32].
- Clean-up: Perform clean-up with ethyl acetate and reconstitute the final extract in an acetone/methanol/isopropanol mixture (40/40/20 v/v/v) [32].
- Internal Standards: Add deuterated internal standards (e.g., d6-Cholesterol, d6-Squalene) at the beginning of extraction to correct for technical variability [32].
Data Acquisition:
- Employ multiple analytical methods for comprehensive coverage [50]:
- Liquid Chromatography-Mass Spectrometry (LC-MS): For abundant lipids (phospholipids, triglycerides, cholesterol esters) and hydrophilic metabolites.
- Gas Chromatography-Mass Spectrometry (GC-MS): For compounds like squalene, cholesterol, saturated fatty acids, and monounsaturated fatty acids [32].
- Targeted Assays: For specific classes such as steroid hormones (e.g., progesterone, estrogen) and lipid mediators (e.g., prostaglandins).
Data Processing and Noise Filtration:
- Process raw data using peak detection and alignment software.
- Apply a noise-filtering algorithm (e.g.,
noisyRor Polly-PeakML) to remove technical noise and retain high-confidence signals [58] [59]. - Perform quantitative normalization and statistical analysis to identify phase-specific metabolic patterns.
Validating Metabolic Rhythmicity in the Menstrual Cycle
After noise reduction, the filtered data must be rigorously analyzed to establish true rhythmicity.
Statistical Analysis:
- Multivariate Analysis: Use Principal Component Analysis (PCA) and Partial Least Squares-Discriminant Analysis (PLS-DA) to visualize global metabolic profile separation between cycle phases or groups (e.g., severe vs. mild menstrual pain) [50].
- Differential Analysis: Generate volcano plots to identify specific metabolites with significant fold-changes and p-values (e.g., |log2FC| > 1 and p < 0.05) [50].
- ROC Analysis: Assess the discriminatory power of candidate biomarkers by calculating the Area Under the Curve (AUC). Ratios of metabolites (e.g., PI(36:2)/BCAA) can enhance accuracy [50].
- Correlation with Clinical Phenotypes: Test for significant linear correlations between metabolite ratios and clinical scores (e.g., Numerical Pain Rating Scale) to validate biological relevance [50].
Downstream Impact on Biological Interpretation
Applying rigorous noise filtration has a profound and immediate consequence on the convergence of predictions in downstream analyses. Studies have demonstrated that filtering out noise leads to more consistent results across different analytical approaches for differential expression calls, enrichment analyses, and the inference of gene regulatory networks [58].
In the context of menstrual cycle research, this translates to a more reliable identification of true amino acid and lipid rhythmicity. For example, without proper noise handling, the inverse relationship between BCAAs and specific phosphatidylinositols across the cycle might be missed or misrepresented. By implementing the described methodologies, researchers can ensure that the biological interpretations and conclusions drawn about cycle-phase specific metabolism are based on robust, high-quality data, ultimately advancing our understanding of female physiology and informing drug development targets for conditions like dysmenorrhea or other menstrual-related disorders.
Standardizing Terminology and Reporting for Cross-Study Comparisons
The investigation of amino acid and lipid rhythmicity across the menstrual cycle represents a frontier in understanding female physiology. Research indicates that the menstrual cycle is characterized by significant metabolic fluctuations, with one comprehensive study identifying 208 of 397 tested metabolites as significantly changed (p < 0.05) across the cycle, 71 of which met a false discovery rate threshold of 0.20 [8] [60] [7]. These rhythmic patterns were particularly evident in neurotransmitter precursors, glutathione metabolism, the urea cycle, and specific micronutrients [8]. A critical finding across multiple studies is that 39 amino acids and derivatives along with 18 lipid species demonstrate decreased concentrations during the luteal phase, potentially indicating an anabolic state during the progesterone peak [8] [7].
Despite these important findings, the field faces substantial challenges in cross-study comparison due to inconsistent terminology, methodology, and reporting practices. Early research often neglected to account for fluctuating hormone profiles or combined eumenorrheic participants with hormonal contraceptive users [49]. Furthermore, studies have employed heterogeneous methodologies for phase verification, including cycle day counting alone, luteinizing hormone testing, or basal body temperature, often without standardization [49]. This methodological heterogeneity complicates the synthesis of evidence and slows progress in understanding the complex interplay between ovarian hormones and metabolic processes.
Table 1: Key Metabolic Fluctuations Across the Menstrual Cycle
| Metabolic Parameter | Direction of Change | Phase of Maximum Change | Magnitude/Examples |
|---|---|---|---|
| Amino Acids | Decrease | Luteal Phase | 39 amino acids and derivatives significantly decreased [8] |
| Specific Lipids | Decrease | Luteal Phase | 18 lipid species significantly decreased [8] |
| Branched-Chain Amino Acids | Varies by condition | Menstrual Phase (in pain studies) | Decreased in severe menstrual pain vs. mild pain [50] |
| Phosphatidylinositols | Increase | Menstrual Phase (in pain studies) | PI(36:2) increased 1.42-fold in severe pain [50] |
| Cholesterol Profiles | Non-linear fluctuation | Throughout cycle | Total, HDL, LDL cholesterol show non-linear patterns [61] |
Standardized Menstrual Cycle Phase Definitions
Phase Terminology and Hormonal Criteria
To enable meaningful cross-study comparisons, researchers must adopt consistent terminology and biochemical criteria for defining menstrual cycle phases. The six-phase model (early follicular, late follicular, ovulation, early luteal, mid-luteal, and late luteal) provides the most granular framework for metabolic studies [49]. Each phase is characterized by distinct hormonal milieus that directly influence metabolic processes:
- Early Follicular/Menstrual Phase: Estradiol levels are low immediately following menses [49].
- Late Follicular Phase: Estradiol levels peak prior to ovulation [49].
- Periovulatory Phase: Characterized by peaks in follicular-stimulating hormone (FSH) and luteinizing hormone (LH) [8] [49].
- Early Luteal Phase: Estradiol drops then rises while progesterone begins to rise [49].
- Mid-Luteal Phase: Both estradiol and progesterone levels peak approximately 7-9 days post-LH surge [49].
- Late Luteal Phase: Estradiol and progesterone levels fall preceding menstruation [49].
The definition of these phases must be based on both hormonal measurements and cycle day tracking to account for inter-individual variability in cycle length, which typically ranges from 21 to 40 days in eumenorrheic women [49] [61].
Quantitative Hormonal Thresholds for Phase Verification
Standardization requires establishing specific hormonal thresholds for phase verification. Research indicates that luteal phase testing should occur 7-9 days post-LH surge detection to ensure adequate progesterone levels and exclude anovulatory participants [49]. The UK Biobank study calculated standardized time within the menstrual cycle using the formula: (days since last menstrual period/days in usual menstrual cycle), where 0 represents cycle start and 1 represents cycle end, with the follicular phase defined as 0.00-0.54 and luteal phase as 0.54-1.00 [61].
Table 2: Recommended Hormonal Thresholds for Phase Verification
| Cycle Phase | Estradiol (pmol/L) | Progesterone (ng/L) | LH/FSH Status | Cycle Day Reference |
|---|---|---|---|---|
| Early Follicular | Low (< threshold) | Low | Baseline | Day 1-5 (varies by cycle length) |
| Late Follicular | Peak concentration | Low | Rising | Variable, pre-ovulatory |
| Periovulatory | High | Low | LH/FSH peak | Day 12-16 (typical) |
| Early Luteal | Dips then rises | Rising | Post-ovulatory | Day 15-19 (typical) |
| Mid-Luteal | Peak with P4 | Peak (~7-9 days post-LH) | Post-ovulatory | Day 19-23 (typical) |
| Late Luteal | Falling | Falling | Pre-menstrual | Day 24-28 (typical) |
Diagram 1: Menstrual Cycle Phase Hormonal Transitions
Methodological Standardization for Metabolic Studies
Sample Collection and Processing Protocols
Standardized protocols for sample collection and processing are essential for generating comparable metabolomic data. Recent methodological advances enable comprehensive metabolic profiling from minimal sample volumes. One innovative approach developed a pretreatment procedure requiring only 100 μL of finger-prick blood, facilitating repeated sampling across cycle phases while minimizing participant burden [50]. This method enabled quantification of 692 metabolites, including 23 steroid hormones, 85 lipid mediators, 429 abundant lipids, and 155 hydrophilic metabolites using five different analytical methods [50].
Recommended standardized protocols include:
- Blood Collection: Consistent timing relative to participant wake time, standardized fasting conditions (recommended ≥4 hours [61]), and uniform collection tubes.
- Sample Processing: Immediate centrifugation at standardized speeds and temperatures, aliquoting protocols to prevent freeze-thaw cycles, and consistent storage at -80°C.
- Quality Controls: Incorporation of pooled quality control samples, standardization of hemolysis assessment, and implementation of sample tracking systems.
Analytical Techniques for Metabolomic Profiling
Comprehensive metabolic profiling requires multiple analytical platforms to capture the diverse chemical properties and concentration ranges of metabolites involved in menstrual cycle rhythmicity:
- Liquid Chromatography-Mass Spectrometry (LC-MS): For broad metabolomic and lipidomic profiling [8] [50].
- Gas Chromatography-Mass Spectrometry (GC-MS): For volatile compounds and metabolites requiring derivatization [8].
- High-Performance Liquid Chromatography with Fluorescence Detection (HPLC-FLD): For specific nutrient analysis including B vitamins [8].
- Clinical Chemistry Analyzers: For standard clinical biochemistry parameters (e.g., glucose, triglycerides, cholesterol) [8] [61].
The nested study design employing multiple analytical methods demonstrated that global plasma metabolite composition shows greater variation between menstrual pain severity groups than across menstrual cycle phases [50], highlighting the importance of comprehensive metabolite coverage.
Experimental Protocols and Workflows
Comprehensive Metabolomic Profiling Protocol
Based on methodologies from key studies in the field, the following standardized protocol is recommended for investigating amino acid and lipid rhythmicity across the menstrual cycle:
Participant Selection Criteria:
- Regularly menstruating women (cycle length 21-40 days) [50] [61]
- Absence of gynecological disorders, recent pregnancy (<1 year), or hormonal contraceptive use (<1 year) [50] [61]
- Confirmation of ovulatory cycles via hormonal assessment [49]
Phase Verification and Sampling:
- Track menstrual cycle using self-reported days since last period and usual cycle length [61]
- Confirm phase with serum hormones (estradiol, progesterone, LH, FSH) [49]
- Collect samples at multiple timepoints across all cycle phases
- Standardize collection time relative to wake time and fasting status [61]
Sample Processing and Analysis:
- Process plasma, urine, and serum samples within 2 hours of collection [8]
- Employ targeted metabolomic profiling using LC-MS and GC-MS [8]
- Analyze lipidomics using appropriate extraction methods [8] [50]
- Quantify B vitamins using HPLC-FLD [8]
- Perform clinical chemistries on automated analyzers [8] [61]
Diagram 2: Experimental Workflow for Menstrual Cycle Metabolomics
Data Analysis and Rhythmicity Assessment
Standardized statistical approaches are critical for identifying rhythmic patterns in metabolic data:
- Multiple Testing Correction: Employ false discovery rate (FDR) correction with standardized thresholds (e.g., FDR < 0.20) for high-dimensional metabolomic data [8].
- Rhythmicity Analysis: Use cosinor analysis or similar methods to identify oscillating metabolites [62].
- Phase Contrast Comparisons: Pre-specify key phase comparisons (e.g., luteal-follicular, luteal-menstrual) to identify phase-specific metabolic changes [8].
- Multivariate Statistics: Implement principal component analysis (PCA) and partial least squares-discriminant analysis (PLS-DA) to identify global metabolite profile differences [50].
The UK Biobank study utilized generalized additive models (GAM) to investigate non-linear associations between menstrual cycle phase and metabolites, demonstrating significant non-linear relationships with cholesterol profiles [61].
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Essential Research Reagents and Materials for Menstrual Cycle Metabolomics
| Reagent/Material | Specification | Research Application | Example Use |
|---|---|---|---|
| LC-MS Systems | High-resolution mass spectrometers | Broad metabolomic profiling | Detection of 397+ metabolites across cycle phases [8] |
| GC-MS Systems | With derivatization capability | Analysis of volatile metabolites | Complementary metabolomic coverage [8] |
| HPLC-FLD Systems | With appropriate columns/detectors | Targeted vitamin analysis | B vitamin quantification in plasma [8] |
| Automated Clinical Analyzers | Standardized platforms | Clinical biochemistry panels | Glucose, triglyceride, cholesterol measurement [61] |
| LH Urine Test Kits | Qualitative surge detection | Ovulation timing confirmation | Phase verification alongside serum hormones [8] |
| Standardized Collection Tubes | EDTA, serum separator, etc. | Biospecimen integrity | Consistent plasma/serum separation across sites |
| Hormone Assay Kits | ELISA, RIA, or automated platforms | Estradiol, progesterone quantification | Phase confirmation with established thresholds [49] |
| Stable Isotope Standards | Isotope-labeled metabolites | Quantification accuracy | Internal standards for mass spectrometry [50] |
Reporting Standards for Cross-Study Comparison
Minimum Reporting Requirements
To enable effective meta-analyses and cross-study comparisons, researchers should include these essential elements in all publications:
- Participant Characteristics: Age, BMI, menstrual cycle characteristics (length, regularity), gynecological history, hormonal contraceptive history, and physical activity levels [49] [61].
- Phase Verification Methods: Specific hormonal thresholds used for phase classification, timing of sample collection relative to LH surge or cycle day, and assay specifications for hormone measurements [49].
- Analytical Methodology: Detailed description of metabolomic platforms, chromatography conditions, mass spectrometry parameters, and quality control procedures.
- Statistical Methods: Clear description of multiple testing correction approaches, rhythmicity analysis methods, and effect size calculations.
Data Sharing and Metadata Standards
Beyond publication requirements, standardized data sharing practices will accelerate progress in the field:
- Public Data Repositories: Deposit raw and processed metabolomic data in public repositories with standardized metadata.
- Hormonal Data: Include individual participant hormone levels rather than only phase classifications.
- Cycle Timing Information: Report exact days since last menstrual period rather than only phase categories.
- Analysis Code: Share statistical analysis scripts for rhythmicity assessment and data processing.
The highly individual nature of metabolic responses to menstrual cycle phases [62] underscores the importance of collecting and reporting individual-level data to enable personalized approaches in future research and clinical applications.
Standardizing terminology, methodology, and reporting practices in menstrual cycle metabolomics research is not merely an academic exercise—it is a fundamental requirement for advancing our understanding of female physiology and developing evidence-based, personalized approaches to women's health. The consistent observation that amino acids and specific lipid species decrease during the luteal phase [8] [7], and the recent discovery that metabolite ratios can discriminate menstrual pain severity [50], highlight the potential of this research area to transform women's healthcare. By adopting the standardized frameworks outlined in this document, researchers can generate comparable data that will accelerate discovery and ultimately lead to novel nutritional, therapeutic, and diagnostic strategies tailored to the rhythmic nature of female physiology.
Biomarker Validation and Comparative Analysis: From Discovery to Clinical Utility
The pursuit of objective molecular markers for subjective physiological experiences represents a frontier in clinical diagnostics. This whitepaper details the technical validation of paired metabolite biomarkers—branched-chain amino acids (BCAAs) and specific phosphatidylinositols (PIs)—for assessing menstrual pain severity. Grounded in a broader research thesis exploring amino acid-lipid rhythmicity across the menstrual cycle, we present a comprehensive methodological framework for biomarker quantification, ratio analysis, and diagnostic performance evaluation via Receiver Operating Characteristic (ROC) curves. The documented approach achieves exceptional discrimination accuracy (AUC up to 0.990), demonstrating the profound potential of metabolite pairs in objective pain assessment. This guide provides researchers, scientists, and drug development professionals with detailed experimental protocols and analytical workflows for replicating and expanding upon these findings.
The regulation of metabolic pathways exhibits profound diurnal and cyclical variation, influenced by genetic, nutritional, and hormonal pacemakers [63] [64]. The broader thesis of our research posits that the interplay between amino acid and lipid metabolism rhythms—termed "amino acid-lipid rhythmicity"—offers a rich source of biomarkers for cyclical physiological states, particularly those tied to the menstrual cycle. Peripheral tissue rhythms, while synchronized with the central circadian clock, are independently influenced by local cues, including hormonal fluctuations [63]. Simultaneously, genetic variations interact with these environmental and hormonal pacemakers, remodeling enhancer-promoter interactions and ultimately shaping an individual's metabolic landscape [63].
Within this complex framework, BCAAs (valine, leucine, isoleucine) and phosphatidylinositols (PIs) emerge as molecules of particular interest. BCAAs are not only essential nutrients but also signaling molecules whose circulating levels are linked to inflammatory states and metabolic health [65]. PIs are fundamental structural and signaling lipids, with specific species like PI(36:2) implicated in various disease pathways [66] [67]. The inverse relationship between these two metabolite classes in the context of menstrual pain suggests a disruption in their coordinated rhythmicity, providing a quantifiable molecular signature for an otherwise subjective experience.
Detailed Experimental Protocols
Study Population and Sample Collection
Participant Recruitment and Group Stratification
- Inclusion Criteria: Enroll healthy, premenopausal women (e.g., age 20-40) with regular menstrual cycles (25-38 days) and confirmed absence of gynecological disorders via recent examination [5].
- Pain Assessment: Administer a standardized pain rating scale, such as the Numerical Pain Rating Scale (NPRS), during the menstrual phase. Stratify participants into severity groups based on a predefined threshold (e.g., severe pain: NPRS ≥ 4; mild pain: NPRS < 4) [5].
- Ethical Considerations: Obtain informed consent and secure approval from an institutional review board (IRB). Ensure participants are explicitly informed of their right to withdraw from the study without penalty [5] [68].
Minimally Invasive Blood Collection and Processing
- Collection Protocol: Collect finger-prick blood samples (approximately 100 µL) across multiple phases of the menstrual cycle (menstrual, follicular, luteal) to capture cyclical metabolic variations [5].
- Plasma Separation: Centrifuge blood samples promptly after collection to separate plasma. Store aliquots at -80°C until analysis to prevent metabolite degradation [5].
Metabolomic Profiling and Quantification
A comprehensive, multi-platform metabolomic approach is critical for capturing metabolites with diverse chemical properties.
Hydrophilic Metabolite Analysis (BCAAs)
- Platform: Employ a combination of Gas Chromatography-Mass Spectrometry (GC-MS) and Liquid Chromatography-Mass Spectrometry (LC-MS).
- Sample Derivatization (for GC-MS): For the aqueous phase of sample extracts, use a two-step derivatization process. First, add methoxamine in pyridine and shake at 30°C for 1.5 hours. Then, add N-Methyl-N-(trimethylsilyl)trifluoroacetamide (MSTFA) with fatty acid methyl esters (FAMEs) as internal standards and shake at 37°C for 30 minutes [64].
- GC-MS Analysis: Inject the derivatized sample onto a GC system (e.g., 7890A GC) coupled to a time-of-flight (TOF) mass spectrometer. Use a high-quality column (e.g., RESTEK RTX-5SIL MS) and a temperature ramp program (e.g., hold at 50°C for 1 min, ramp to 330°C at 20°C/min, hold for 5 min) [64].
- LC-MS Analysis: Inject the underivatized aqueous phase onto a suitable LC-MS system for complementary quantification.
Lipidomic Analysis (Phosphatidylinositols)
- Lipid Extraction: Use a Matyash-type liquid-liquid extraction buffer containing methyl-tert-butyl ether (MTBE), methanol, and water. The organic (upper) phase containing lipids is dried down and reconstituted for LC injection [64].
- LC-MS Analysis: Utilize reverse-phase liquid chromatography coupled to a high-resolution mass spectrometer to separate and quantify individual PI species. Identify specific PIs like PI(36:2) based on their mass-to-charge ratio and retention time.
Targeted Bioactive Molecule Analysis
- Steroid Hormones and Lipid Mediators: Incorporate additional targeted LC-MS/MS methods to quantify steroid hormones (e.g., progesterone, 17β-estradiol) and lipid mediators (e.g., prostaglandins) to validate cycle phases and provide additional context [5].
Data Processing and Statistical Analysis
Metabolite Quantification and Normalization
- Peak Integration: Integrate raw chromatographic peaks using instrument software.
- Normalization: Normalize metabolite peak areas using internal standards to correct for technical variation. Further normalization to total protein concentration or a systematic quality control (QC) sample pool may be applied.
Multivariate and Univariate Statistics
- Multivariate Analysis: Perform Principal Component Analysis (PCA) and Partial Least Squares-Discriminant Analysis (PLS-DA) on the global metabolite profile to visualize group separations and identify metabolites contributing most to the variance [5].
- Differential Analysis: Generate volcano plots to identify metabolites with statistically significant fold-changes (e.g., >1.2) and p-values (e.g., <0.015) between severe and mild pain groups [5].
Biomarker Ratio Calculation and ROC Analysis
- Ratio Formulation: Calculate the ratios of promising biomarker candidates, specifically PI(36:2) to each individual BCAA (valine, leucine, isoleucine) and to the total BCAA level [5].
- ROC Curve Generation: Perform ROC analysis on both individual metabolites and their ratios to evaluate diagnostic performance. Calculate the Area Under the Curve (AUC) to quantify the ability of each marker to discriminate between pain severity groups [5].
- Correlation with Pain Score: Conduct linear regression analysis to assess the correlation between the identified biomarker ratios and the NPRS scores across all menstrual cycle phases [5].
Figure 1: Experimental workflow for biomarker validation.
Key Findings and Data Presentation
Metabolite Identification and Diagnostic Performance
The application of the above protocols identified specific PIs and BCAAs as significantly altered in individuals with severe menstrual pain.
Table 1: Performance of Individual Metabolites in Discriminating Menstrual Pain Severity
| Metabolite | Fold Change (Severe/Mild) | AUC | Sensitivity/Specificity (%) |
|---|---|---|---|
| PI(36:2) | 1.42 ↑ | 0.906 | - |
| Valine | 0.80 ↓ | 0.865 | - |
| Isoleucine | 0.80 ↓ | 0.865 | - |
| Leucine | 0.80 ↓ | 0.865 | - |
| PI(38:4) | >1.2 ↑ | >0.8 | - |
| PI(38:3) | >1.2 ↑ | >0.8 | - |
| 4-Hydroxyproline | >1.2 ↑ | >0.8 | - |
The most significant finding was the enhanced diagnostic power achieved by using metabolite ratios.
Table 2: Diagnostic Performance of PI(36:2)-BCAA Ratios (Menstrual Phase)
| Biomarker Ratio | Area Under Curve (AUC) |
|---|---|
| PI(36:2) / Total BCAA | 0.990 |
| PI(36:2) / Leucine | 0.979 |
| PI(36:2) / Isoleucine | 0.969 |
| PI(36:2) / Valine | 0.969 |
Crucially, these biomarker ratios maintained high discrimination accuracy across all phases of the menstrual cycle (AUC > 0.8 in follicular and luteal phases) and showed a significant positive linear correlation with NPRS scores, enabling prediction and monitoring of pain severity beyond the immediate menstrual period [5].
Integration with Broader Thesis: Metabolic Rhythmicity
The inverse relationship between PIs and BCAAs suggests a disruption in their coordinated metabolic rhythms during painful menstruation. This aligns with the broader concept that genetic and environmental factors can remodel metabolic pathways. Research shows that genetic variants within regulatory elements interact with nutritional signals to control diurnal enhancer-promoter dynamics and liver lipid metabolism [63]. While not yet fully elucidated in the endometrium, similar mechanisms likely govern the rhythmicity of amino acid and lipid metabolism in the uterus. The identified biomarker pairs may thus be reflecting a fundamental dysregulation of this metabolic clockwork.
Signaling Pathways and Molecular Context
The biological plausibility of the BCAA-PI relationship is supported by their interconnected roles in cellular signaling and metabolism.
BCAA Metabolism and Signaling: BCAAs are essential amino acids whose catabolism is tightly regulated. High BCAA levels can activate the mTOR signaling pathway, a central regulator of cell growth and proliferation, and are strongly associated with insulin resistance and inflammatory states [65] [69]. A decrease in BCAA levels, as observed in severe dysmenorrhea, may reflect either increased catabolism or altered flux through these signaling pathways.
Phosphatidylinositol Signaling: PIs are minor membrane phospholipids that serve as precursors to critical signaling molecules. Through site-specific phosphorylation by kinases like PI3K, they generate lipid second messengers (e.g., PIP2, PIP3) that regulate a vast array of cellular processes, including cytoskeletal organization, membrane trafficking, and cell survival [66] [67]. The specific elevation of PI(36:2) could indicate altered activity of these kinase/phosphatase networks in the context of uterine inflammation and pain.
Figure 2: Proposed signaling pathways for BCAA and PI interactions.
The convergence of BCAA and PI metabolism on inflammatory pathways provides a plausible mechanistic link to the pathophysiology of menstrual pain, which is characterized by elevated levels of inflammatory prostaglandins and uterine inflammation [5].
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents and Materials for Experimental Replication
| Item | Function / Specification | Example / Note |
|---|---|---|
| Lancets | Finger-prick blood collection | Ensure safety and standardized depth. |
| EDTA/Lithium Heparin Microtainers | Plasma collection and anticoagulation | For 100 µL blood volumes. |
| MTBE/MeOH/H2O Solution | Lipid extraction (Matyash method) | Organic phase for PI analysis. |
| MSTFA with 1% TMCS | Derivatization agent for GC-MS | For silylation of BCAAs. |
| Pyridine & Methoxamine HCl | Derivatization for GC-MS | Protects carbonyl groups. |
| Deuterated Internal Standards | e.g., d₃-Leucine, d₅-PI(34:1) | Essential for MS quantification. |
| RESTEK RTX-5SIL MS Column | GC separation of derivatized metabolites | 30m length, 0.25mm i.d. |
| C18 Reverse-Phase LC Column | UPLC/HPLC separation of PIs | 1.7-1.8 µm particle size. |
| Quality Control (QC) Pooled Plasma | Monitor instrument performance | Pool from all study samples. |
This technical guide validates the ratio of PI(36:2) to BCAAs as a highly accurate biomarker pair for objectively assessing menstrual pain severity. The methodological framework—from minimally invasive sampling and multi-platform metabolomics to rigorous ROC analysis—provides a robust blueprint for researchers. The findings are powerfully contextualized within a broader thesis of amino acid-lipid rhythmicity, suggesting that the dysregulation of these interconnected metabolic pathways underlies the subjective experience of pain. Future work should focus on validating these ratios in larger, more diverse cohorts and elucidating the precise molecular mechanisms that link BCAA-PI rhythmicity to uterine physiology and pain perception, ultimately paving the way for novel diagnostic tools and targeted therapeutic interventions.
The human body is governed by complex, interconnected physiological rhythms, from the rapid-fire rhythm of the heartbeat to the daily oscillations of the circadian clock. The menstrual cycle represents another essential, yet understudied, life rhythm, orchestrated by the intricate interplay of hormones like progesterone, estradiol, follicular stimulating hormone (FSH), and luteinizing hormone (LH) [1]. While the rhythmicity of sex hormones is well-documented, its profound influence on the cellular metabolome is only beginning to be understood. Metabolomics, the comprehensive analysis of small molecule metabolites, provides a dynamic snapshot of physiological status, capturing the influence of genetics, environment, and lifestyle [70]. This technical guide explores how comparative metabolomics unveils the distinct metabolic patterns of the menstrual cycle, framing these findings within a broader thesis on amino acid and lipid rhythmicity. For researchers and drug development professionals, this synthesis aims to illuminate the unique metabolic landscape of the female cycle, contrast it with other physiological states, and provide a foundation for novel, cycle-aware diagnostic and therapeutic strategies.
Metabolic Rhythmicity of the Menstrual Cycle
Phase-Dependent Fluctuations in Amino Acids and Lipids
Advanced metabolomic profiling of healthy, premenopausal women has revealed that the menstrual cycle is far from metabolically static. A study analyzing 397 metabolites in plasma and urine across five cycle phases (Menstrual, Follicular, Periovulatory, Luteal, Premenstrual) found that 208 were significantly changed (p < 0.05), with 71 meeting a False Discovery Rate (FDR) threshold of 0.20 [1]. The most striking changes were observed in amino acids and lipid species, showing a clear rhythmicity.
Amino Acids and Biogenic Amines: Of 54 amino acids and derivatives detected, 48 showed significant variation across cycle phases [1]. A consistent pattern emerged, with a significant decrease in plasma levels during the luteal phase. For instance, ornithine, arginine, alanine, glycine, methionine, and proline were significantly lower in the luteal phase compared to all other phases [1]. After multiple testing correction, 37 amines remained significantly reduced in the Luteal-Menstrual (L-M) contrast [1]. This widespread reduction may indicate a state of increased nitrogen utilization and protein turnover during the post-ovulatory period, potentially representing an anabolic state driven by the progesterone peak [1].
Lipid Species: The luteal phase is also characterized by a significant reduction in specific phospholipids. Out of 139 lipid species analyzed, 57 showed significant variation, with 38% consistently decreasing in the luteal phase [1]. Seventeen lipid species, including 6 lysophosphatidylcholines (LPCs), 10 phosphatidylcholines (PCs), and 1 lysophosphatidylethanolamine (LPE), met the FDR threshold for the Luteal-Follicular (L-F) contrast [1]. The metabolite LPE 22:6 was particularly notable, showing significance in four out of five phase contrasts [1].
Table 1: Significant Metabolite Changes Across the Menstrual Cycle (FDR < 0.20)
| Metabolite Class | Specific Metabolites | Direction of Change | Key Phase Contrast |
|---|---|---|---|
| Amino Acids & Biogenic Amines | Ornithine, Arginine, Alanine, Glycine, Methionine, Proline, Threonine, Serine | ↓ in Luteal Phase | Luteal vs. Menstrual (L-M), Luteal vs. Follicular (L-F) |
| Phospholipids | LPCs (6 species), PCs (10 species), LPE 22:6 | ↓ in Luteal Phase | Luteal vs. Follicular (L-F) |
| Vitamins & Cofactors | 25-OH Vitamin D, 4-Pyridoxic Acid (Vitamin B6 metabolite) | ↑ in Menstrual Phase | Luteal vs. Menstrual (L-M), Periovulatory vs. Menstrual (O-M) |
Menstrual Pain as a Model of Dysregulated Rhythmicity
The natural metabolic rhythmicity of a healthy cycle can be perturbed by conditions like dysmenorrhea (menstrual pain). A 2025 metabolomics study identified potential plasma biomarkers for pain severity, highlighting the interplay between lipids and amino acids [5]. The study, which quantified 692 metabolites from finger-prick blood, found that the global plasma metabolite profile was more strongly associated with pain severity than with the menstrual cycle phase itself [5].
Key biomarkers identified include:
- Phosphatidylinositol (PI(36:2)): Significantly increased in the severe pain group (1.42-fold).
- Branched-Chain Amino Acids (BCAAs - leucine, isoleucine, valine): Significantly decreased in the severe pain group (to ~0.8-fold).
Notably, the ratio of PI(36:2) to total BCAAs showed an exceptional ability to discriminate between severe and mild pain groups, with an Area Under the Curve (AUC) of 0.990 in Receiver Operating Characteristic (ROC) analysis. This ratio also demonstrated a significant positive linear correlation with the Numerical Pain Rating Scale (NPRS) across all menstrual cycle phases, suggesting its utility as an objective, phase-agnostic biomarker for menstrual pain [5].
Contrasting Menstrual and Pathological Metabolic States
Polycystic Ovary Syndrome (PCOS)
Comparing the rhythmic metabolome of a healthy menstrual cycle with the metabolic signature of Polycystic Ovary Syndrome (PCOS) reveals stark contrasts. PCOS is characterized by hormonal imbalances and metabolic complications, and its metabolomic profile is further influenced by environmental factors [70].
A comparative study of urban and rural women with PCOS using LC-MS/MS showed distinct metabolic disruptions:
- Rural PCOS: Exhibited higher levels of lipid-related metabolites, particularly Palmitone, suggesting influences from a diet rich in natural fats [70].
- Urban PCOS: Showed more pronounced alterations in carbohydrate and nucleotide metabolism pathways, likely linked to the consumption of processed foods [70].
Furthermore, a study analyzing follicular fluid metabolomes identified specific metabolites that powerfully discriminate PCOS from other infertility phenotypes like Poor Ovarian Response (POR) [71]. Trehalose-6-phosphate, taurocholate, and N,N-dimethylglycine emerged as the most significantly altered metabolites. PCOS patients exhibited reduced levels of taurocholate, mycalemide, and trehalose-6-phosphate compared to other groups [71]. These findings point to fundamental disruptions in bile acid metabolism and glycosylation processes in PCOS, which stand in contrast to the cyclical, rhythmic changes seen in healthy premenopausal women.
Table 2: Contrasting Metabolic Signatures: Healthy Cycle vs. PCOS
| Aspect | Healthy Menstrual Cycle | Polycystic Ovary Syndrome (PCOS) |
|---|---|---|
| Overall Pattern | Rhythmic, phase-dependent fluctuations | Stable, dysregulated metabolic state |
| Key Amino Acids | Cyclic decrease in most AAs during luteal phase | Phenotype-specific alterations (e.g., N,N-dimethylglycine) |
| Key Lipids | Cyclic decrease in specific phospholipids (PCs, LPCs) in luteal phase | Elevated lipid metabolites (rural); altered carb/nucleotide metabolism (urban) |
| Discriminatory Metabolites | Patterns of ~70 metabolites across cycle | Taurocholate, Trehalose-6-phosphate, N,N-dimethylglycine |
| Environmental Influence | Consistent rhythmic pattern | Signature varies significantly with urban/rural environment |
Essential Methodologies for Menstrual Cycle Metabolomics
Experimental Workflow for Serum/Plasma Metabolomics
Robust metabolomic profiling requires standardized protocols from sample collection to data analysis. The following workflow is common to several cited studies [1] [70] [5].
Title: Metabolomics Experimental Workflow
1. Sample Collection & Preparation:
- Timing: Blood samples should be collected during specific, verified menstrual cycle phases (e.g., early follicular phase: days 2-5) after an overnight fast [70] [71].
- Processing: Collect blood into serum separator tubes. Centrifuge at 2,500-3,000 rpm for 15 minutes at 4°C. Aliquot the supernatant (serum/plasma) and store immediately at -80°C until analysis [70] [71].
2. Metabolite Extraction:
- Protein Precipitation: Precisely measure 100-300 µL of serum. Add 400-600 µL of chilled methanol or acetonitrile (LC-MS grade) to precipitate proteins [5] [71].
- Incubation & Centrifugation: Vortex vigorously, then incubate at -20°C for 15 minutes. Centrifuge at 10,000-12,000 g for 10-15 minutes at 4°C to pellet proteins [5] [71].
- Supernatant Collection: Carefully transfer the clear supernatant to a new vial. Vacuum-dry or evaporate under nitrogen gas, then reconstitute in a solvent compatible with the analytical instrument (e.g., acetonitrile/water mixture). Filter through a 0.22 µm syringe filter prior to injection [70] [5].
3. Liquid Chromatography-Mass Spectrometry (LC-MS) Analysis:
- Chromatography: Use reverse-phase chromatography (e.g., C18 column). A typical mobile phase consists of (A) water with 0.1% formic acid and (B) methanol or acetonitrile, with a gradient elution from 5% B to 95% B over 15-20 minutes [70] [71].
- Mass Spectrometry: Perform data acquisition using high-resolution mass spectrometers (e.g., Q-Exactive Plus Orbitrap) in full-scan mode, with both positive and negative electrospray ionization (ESI) to maximize metabolite coverage [5] [71].
4. Data Processing and Bioinformatics:
- Conversion: Convert raw data files to open formats (mzXML, .mgf) using software like ProteoWizard [71].
- Peak Alignment & Annotation: Use platforms like MS-DIAL or XCMS for peak picking, alignment, and deconvolution. Annotate metabolites by matching accurate mass and fragmentation spectra against databases such as HMDB, MassBank, and MoNA [5] [71].
- Statistical Analysis: Employ multivariate statistics like Principal Component Analysis (PCA) and Orthogonal Projections to Latent Structures-Discriminant Analysis (OPLS-DA) to identify group separations. Perform univariate tests (t-tests, ANOVA) with False Discovery Rate (FDR) correction. Conduct pathway analysis using KEGG or MetaboAnalyst [70] [5] [71].
The Scientist's Toolkit: Key Research Reagents and Solutions
Table 3: Essential Reagents and Materials for Menstrual Cycle Metabolomics
| Item | Function / Purpose | Example / Specification |
|---|---|---|
| LC-MS Grade Solvents | Mobile phase preparation; minimizes background noise and ion suppression. | J.T. Baker Acetonitrile, Water, Methanol; Formic Acid [70] |
| Protein Precipitation Solvents | Deproteinization of serum/plasma samples for metabolite extraction. | Chilled Methanol or Acetonitrile [5] [71] |
| Standard Metabolite Libraries | Metabolite identification by matching mass and fragmentation patterns. | Human Metabolome Database (HMDB), MassBank, MoNA [5] [71] |
| Quality Control (QC) Pool | Monitors instrument stability and performance throughout the analytical run. | Pooled sample from all study aliquots, analyzed intermittently [70] |
| Chromatography Column | Separation of complex metabolite mixtures prior to mass spectrometry. | Fortis C18 column (3 µm, 150 x 2.1 mm) [71] |
| Syringe Filter | Removal of particulate matter from reconstituted samples to protect the LC system. | 0.22 µm Nylon or Cellulose Acetate [70] |
Visualization of Multi-Omics Data in Metabolic Pathways
Integrating metabolomic data with other omics layers (e.g., transcriptomics, proteomics) provides a more systems-level understanding of menstrual cycle rhythmicity. Specialized tools enable the simultaneous visualization of multiple datasets on organism-scale metabolic network diagrams.
Key Capabilities of Multi-Omics Visualization Tools [72]:
- Multiple Visual Channels: Different omics datasets can be painted onto the same metabolic chart. For example, transcriptomics data can be represented by reaction arrow color, proteomics data by arrow thickness, and metabolomics data by metabolite node color.
- Interactivity and Zooming: Tools support semantic zooming, revealing more detail as the user zooms in. They also allow for the graphing of data values for user-designated reactions or metabolites.
- Dynamic Data Representation: For time-series data, such as those collected across menstrual cycle phases, the tools can display changes in an animated fashion, visually tracking metabolic shifts from the follicular to the luteal phase.
Title: Hormonal Regulation of Metabolism
Discussion and Future Directions
This synthesis of comparative metabolomics underscores that the healthy human menstrual cycle is a state of predictable metabolic rhythmicity, particularly for amino acids and complex lipids. The recurrent decrease in these metabolites during the luteal phase may represent a window of vulnerability, where the system is stressed by high progesterone and more susceptible to symptoms like those of Premenstrual Syndrome (PMS) or Premenstrual Dysphoric Disorder (PMDD) [1]. The contrast with the stable, yet dysregulated, metabolomes of conditions like PCOS highlights the importance of a cyclical framework for interpreting female metabolic data. Furthermore, the finding that symptom burden (e.g., pain) has a more significant impact on certain metabolic readouts (like sleep quality and BCAA/PI ratios) than the cycle phase itself [5] [19] argues for a research and clinical model that integrates both hormonal phase and subjective experience.
For the field of amino acid and lipid rhythmicity, these findings open several research pathways:
- Mechanistic Studies: Determine the hormonal drivers (progesterone vs. estrogen) of the observed luteal-phase decreases in amino acids and lipids using in vitro models.
- Nutritional Interventions: Develop phase-specific nutritional strategies to test if supplementing depleting metabolites (e.g., specific amino acids) in the luteal phase can alleviate symptoms and restore metabolic balance.
- Drug Timing (Chronotherapy): Investigate whether the efficacy or pharmacokinetics of drugs metabolized through affected pathways (e.g., glutathione metabolism, urea cycle) vary across the menstrual cycle.
- Multi-Omics Integration: Combine metabolomic data with transcriptomic and proteomic analyses from the same subjects to build a more complete model of the regulatory network governing menstrual cycle rhythmicity.
In conclusion, moving beyond a static view of female metabolism to embrace its inherent rhythmicity is not merely a niche consideration but a fundamental requirement for precision medicine. Incorporating cycle phase and symptom tracking into study designs and clinical assessments will unlock deeper biological insights and pave the way for more effective, personalized health solutions for women.
The investigation of rhythmic fluctuations in amino acids and lipids across the menstrual cycle represents a frontier in personalized medicine, particularly for understanding female-specific metabolic physiology. While numerous studies have documented metabolic changes throughout the cycle, cross-population validation remains essential for distinguishing universal physiological patterns from population-specific characteristics. This review synthesizes findings from Chinese, European, and Japanese cohorts to establish a robust, validated framework for amino acid and lipid rhythmicity during the menstrual cycle, providing researchers and drug development professionals with methodological standards and comparative insights.
Comparative Analysis of Cohort Findings
Table 1: Overview of menstrual cycle metabolite studies across populations
| Population | Cohort Size | Key Metabolites Studied | Primary Findings | Reference |
|---|---|---|---|---|
| Chinese | 38 women | Sebum lipids (triglycerides, wax esters, squalene, FFAs) | Significant fluctuation of facial sebogenesis during menstrual cycle; HS group showed more sebum lipids than LS group | [32] |
| European (UK Biobank) | 8,694 women | Glucose, triglyceride, total cholesterol, HDL, LDL | Non-linear associations between menstrual cycle phase and cholesterol profiles | [73] |
| Japanese | 20 women | BCAAs, phosphatidylinositols, steroid hormones, lipid mediators | PI(36:2)/BCAA ratio effectively discriminates menstrual pain severity | [50] |
Quantitative Metabolite Changes Across Populations
Table 2: Metabolite fluctuations across menstrual cycle phases
| Metabolite Class | Specific Metabolites | Chinese Cohort Findings | European Cohort Findings | Japanese Cohort Findings |
|---|---|---|---|---|
| Sebum Lipids | Triglycerides, Wax Esters, Squalene | 30-50% TGs, 20-30% WEs, 12-20% squalene in sebum composition; percentage of squalene higher in females | Not assessed | Not assessed |
| Blood Lipids | Total, HDL, and LDL cholesterol | Not assessed | Significant non-linear associations with menstrual cycle (p<0.001 for total and HDL, p=0.012 for LDL) | Not assessed |
| Amino Acids | Branched-chain amino acids (BCAAs) | Not assessed | Not assessed | Decreased to approximately 0.8-fold in severe pain group; effective discriminators of pain severity (AUC: 0.865) |
| Phospholipids | Phosphatidylinositols | Not assessed | Not assessed | PI(36:2) increased 1.42-fold in severe pain group; highest discrimination accuracy (AUC: 0.906) |
| Metabolite Ratios | PI(36:2)/BCAA | Not assessed | Not assessed | Most effective discrimination (AUC: 0.990); positive linear correlation with NPRS |
Methodological Approaches for Cross-Population Studies
Standardized Experimental Protocols
Participant Selection and Menstrual Cycle Phase Determination
The Chinese cohort study implemented rigorous inclusion criteria focusing on women aged 24-29 years with self-declared cycle duration of 26-30 days [32]. Participants recorded bleeding days for two months preceding sample collection, with ovulation monitored using daily ovulation tests starting from day 8 until positive results were observed. The study categorized subjects into low sebum (LS, ≤70 μg/cm²) and high sebum (HS, ≥150 μg/cm²) groups based on sebum accumulation overnight on foreheads using a Sebumeter (Courage and Khazaka, Cologne, Germany) [32].
The UK Biobank study utilized a substantially larger cohort (n=8,694) of regularly menstruating women aged 37-73 years [73]. Menstrual cycle phase was assessed using self-reported answers to "How many days since your last menstrual period?" and "How many days is your usual menstrual cycle?" Standardized time within the menstrual cycle was calculated relative to each participant using the formula: (days since last menstrual period/days in usual menstrual cycle), with 0 representing cycle start and 1 representing cycle end [73].
The Japanese study enrolled 20 healthy women with regular menstrual cycles (25-38 days) and minimal variation (≤6 days) [50]. Recent gynecological examinations confirmed absence of disorders. Participants were divided into severe pain (NPRS≥4, n=8) and mild pain (NPRS<4, n=12) groups based on numerical pain rating scale scores.
Sample Collection and Processing Protocols
Sebum Sampling (Chinese Cohort): After face washing with standardized cleanser, isopropanol wipes removed surface sebum (confirmed using Sebumeter reading under 10 μg/cm²) [32]. Cuderm S100 Sebutape patches (28.6 mm × 19.1 mm) were applied to left and right sides of forehead central line and cheeks for 30 minutes. Cleaned forceps peeled off patches, which were folded into disposable tubes and stored at -80°C freezer until analysis. Sebum was collected every 2 days, with samples corresponding to follicular phase (FP), ovulatory phase (OP), early luteal phase (ELP), and late luteal phase (LLP) processed for sebum lipid analyses.
Blood Sampling (Japanese Cohort): The Japanese study developed a minimally invasive protocol using only 100 μL of finger-prick blood, enabling evaluation across multiple menstrual cycle phases [50]. This approach facilitated comprehensive metabolite profiling while reducing participant burden, potentially increasing compliance in longitudinal study designs.
Blood Sampling (UK Biobank): The UK Biobank followed standardized blood sampling protocols with serum concentrations of metabolites assessed using the AU5800 (Beckman Coulter, CA, USA) [73]. The triglyceride and glucose index (TyG) was calculated as (Ln[(triglyceride mg/dL) × fasting glucose mg/dL)/2]).
Analytical Techniques for Metabolite Quantification
Lipidomics Approaches
The Chinese cohort employed both gas chromatography and liquid chromatography mass spectrometry techniques for comprehensive sebum lipid analysis [32]. For GCMS analysis, 10 μL of dissolved sebum extract was dried under nitrogen and derivatized with 50 μL BSTFA added with 1% trimethylchlorosilane (TCMS) in pyridine to generate trimethylsilyl (TMS) derivatives [32]. Quantitative analysis targeted squalene, cholesterol, saturated FAs (SFAs), monounsaturated FAs (MUFAs), and FAOHs.
The Japanese study implemented five different analytical methods to quantify both water-soluble and organic solvent-soluble small molecules across a broad concentration range [50]. This comprehensive profiling detected 692 metabolites, including 23 steroid hormones, 85 lipid mediators, 429 abundant lipids, and 155 hydrophilic metabolites, providing unprecedented coverage of metabolic fluctuations.
Data Analysis and Statistical Approaches
The UK Biobank study utilized generalized additive models (GAM) to investigate non-linear associations between menstrual cycle phase and outcome variables [73]. Moderator analysis explored whether anthropometric, fitness, physical activity, or inflammatory markers modified associations between menstrual cycle phase and metabolites.
The Japanese study employed multivariate analyses including principal component analysis (PCA) and partial least squares discriminant analysis (PLS-DA) to examine global plasma metabolite composition differences [50]. Receiver operating characteristic (ROC) analysis evaluated the efficacy of metabolites as biomarkers for severe menstrual pain, with area under the curve (AUC) calculations determining discriminatory power.
Signaling Pathways and Metabolic Networks
Diagram 1: Integrated metabolic pathways in menstrual cycle rhythmicity
The molecular interplay between amino acid and lipid metabolism across the menstrual cycle involves complex regulatory networks. Hormonal fluctuations drive metabolic shifts through multiple signaling pathways, including mTORC1 activation which responds to both hormonal changes and amino acid availability [74]. The skeletal muscle circadian clock transcriptionally programs both lipid and amino acid metabolism, with BMAL1 promoting diurnal cycles of neutral lipid storage while coordinately inhibiting lipid and protein catabolism [75].
BCAA metabolism emerges as a crucial node, with leucine specifically activating mTORC1 through receptor-mediated pathways [74]. The Japanese cohort identified the PI(36:2)/BCAA ratio as particularly significant, demonstrating consistent discrimination accuracy across all menstrual cycle phases with significant positive linear correlation with pain scores (NPRS) [50]. This suggests integrated metabolic dysregulation rather than isolated pathway involvement.
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Key research reagents and materials for menstrual cycle metabolite studies
| Category | Specific Items | Application/Function | Examples from Studies |
|---|---|---|---|
| Sample Collection | Sebutape patches (Cuderm S100) | Sebum absorption from skin surface | Chinese cohort: 28.6 mm × 19.1 mm patches applied to forehead and cheeks [32] |
| Finger-prick blood collection devices | Minimally invasive blood sampling | Japanese cohort: 100 μL finger-prick blood for comprehensive metabolite profiling [50] | |
| Analytical Instruments | GC-MS systems | Separation and quantification of sebum lipids | Chinese cohort: GC 7890A coupled to MS 5975 VL analyzer (Agilent Technologies) [32] |
| LC-MS systems | Comprehensive lipidomics and metabolomics | Japanese cohort: Five analytical methods for 692 metabolites [50] | |
| Sebumeter (Courage & Khazaka) | Quantitative sebum measurement | Chinese cohort: Categorization into LS (≤70 μg/cm²) and HS (≥150 μg/cm²) groups [32] | |
| Chemical Reagents | Deuterated internal standards | Quantitative accuracy in mass spectrometry | Chinese cohort: d6Cholesterol, d6Squalene, d17C18:1, d5TG 48:0 [32] |
| Derivatization reagents | Volatilization for GC-MS analysis | Chinese cohort: BSTFA with 1% TCMS in pyridine for TMS derivatives [32] | |
| Antioxidants | Sample integrity preservation | Chinese cohort: BHT (0.025%) in ethanol to prevent oxidation [32] | |
| Hormonal Assays | Salivary hormone tests | Non-invasive hormone monitoring | Elite athlete study: Salivary hormone samples twice weekly [19] |
| Ovulation test kits | Cycle phase confirmation | Chinese cohort: Daily ovulation tests from day 8 until positive results [32] |
Cross-Population Consensus and Divergence
Universal Physiological Patterns
Despite methodological and population differences, consistent themes emerge across studies. The UK Biobank findings demonstrated significant non-linear relationships between menstrual cycle phase and cholesterol profiles, particularly for total cholesterol (p<0.001), HDL (p<0.001), and LDL (p=0.012) [73]. This suggests fundamental endocrine regulation of lipid metabolism transcending ethnic boundaries.
Similarly, the Japanese cohort's identification of BCAA involvement in menstrual symptoms aligns with broader understanding of amino acid metabolism regulation by hormonal fluctuations. The finding that BCAAs decreased to approximately 0.8-fold in the severe pain group while PI(36:2) increased 1.42-fold indicates coordinated metabolic reprogramming rather than isolated changes [50].
Population-Specific Considerations
The Chinese cohort's detailed sebum analysis revealed compositional specifics, including triglycerides (30-50%), wax esters (20-30%), and squalene (12-20%) as major components, with squalene percentage tending to be higher in females [32]. These quantitative benchmarks provide population-specific reference values for future comparative studies.
Methodological differences in sample collection—from sebum tapes in the Chinese cohort to finger-prick blood in the Japanese cohort and venous blood in the UK Biobank—highlight both the diversity of approaches and the need for standardized protocols in cross-population research [32] [73] [50].
Cross-population validation provides crucial insights into universal versus population-specific aspects of amino acid and lipid rhythmicity across the menstrual cycle. The consistency in cholesterol fluctuations in the European cohort and the BCAA-phosphoinositol relationship in the Japanese cohort suggest fundamental metabolic regulatory mechanisms, while quantitative differences in sebum composition highlight potential population-specific characteristics.
Future research should prioritize standardized protocols enabling direct cross-population comparisons, expanded metabolite panels capturing both amino acids and lipids simultaneously, and longitudinal designs tracking individuals across multiple cycles. Integration of multi-omics approaches will further elucidate the complex interplay between hormonal fluctuations and metabolic pathways, advancing both fundamental knowledge and targeted interventions for menstrual cycle-related disorders.
Within the broader investigation of amino acid and lipid rhythmicity across the menstrual cycle, a critical frontier lies in linking these dynamic metabolic signatures to tangible clinical endpoints. Fluctuations in sex hormones regulate sebaceous gland cell lipogenesis and influence the chemical composition of skin surface lipids [32]. Beyond dermatological applications, the rhythmic variation of metabolites offers a powerful lens through which to view and objectively quantify patient-reported outcomes such as pain severity. This whitepaper provides an in-depth technical guide for researchers and drug development professionals on the current methodologies, key biomarkers, and analytical frameworks for connecting specific metabolic patterns to clinical pain and performance measures, with a specific focus on menstrual cycle research.
Metabolic Signatures of Pain Severity
Comprehensive metabolomic profiling is revealing specific plasma metabolites that serve as objective biomarkers for pain severity, effectively translating subjective experience into quantifiable data.
Key Biomarker Pairs for Menstrual Pain
A 2025 metabolomics study utilizing five analytical methods identified distinct metabolite pairs that effectively discriminate between mild and severe menstrual pain. The research, which profiled 692 metabolites from finger-prick blood samples, highlighted branched-chain amino acids (BCAAs) and specific phosphatidylinositols (PIs) as key indicators [50].
Table 1: Metabolite Biomarkers for Menstrual Pain Severity
| Metabolite | Direction in Severe Pain | AUC (Pain Severity Discrimination) | Biological Class |
|---|---|---|---|
| PI(36:2) | ↑ 1.42-fold increase | 0.906 | Phosphatidylinositol Lipid |
| Valine | ↓ ~0.8-fold decrease | 0.865 | Branched-Chain Amino Acid |
| Isoleucine | ↓ ~0.8-fold decrease | 0.865 | Branched-Chain Amino Acid |
| Leucine | ↓ ~0.8-fold decrease | 0.865 | Branched-Chain Amino Acid |
| PI(36:2)/Total BCAA Ratio | Inverse Relationship | 0.990 | Combined Metric |
The most powerful discriminatory performance was achieved not by individual metabolites, but by their ratios. The ratio of PI(36:2) to total BCAAs showed an exceptional area under the curve (AUC) of 0.990 for distinguishing pain severity, and these ratios also demonstrated a significant positive linear correlation with numerical pain rating scale (NPRS) scores across all menstrual cycle phases [50]. This suggests the potential for a robust, phase-agnostic biomarker for objective pain assessment.
Broad Metabolic Profiling for Multidisease Risk
Beyond specific pain conditions, NMR-derived metabolomic profiles demonstrate broad utility in stratifying risk for numerous clinical endpoints. A large-scale study modeling 168 metabolic markers from over 117,000 UK Biobank participants showed that disease-specific "metabolomic states" can inform the risk of 24 common conditions [76].
Table 2: Select Metabolomic State Associations with 10-Year Disease Risk
| Disease Endpoint | Event Rate (Top 10% Metabolomic State) | Event Rate (Bottom 10% Metabolomic State) | Odds Ratio |
|---|---|---|---|
| Type 2 Diabetes | 21.87% | 0.36% | 61.45 |
| Abdominal Aortic Aneurysm | 2.46% | 0.18% | 14.10 |
| Heart Failure | 10.80% | 0.96% | 11.27 |
| All-Cause Dementia | 3.30% | 0.52% | 6.39 |
This approach demonstrates that a single metabolomic assay can provide predictive information for a range of endpoints, from metabolic and vascular diseases to neurological conditions, often outperforming or adding value to conventional clinical predictors [76].
Experimental Protocols and Methodologies
Robust experimental design is paramount for reliably capturing metabolic fluctuations and linking them to clinical endpoints.
Comprehensive Metabolomic Workflow for Menstrual Pain
The identified biomarker pairs for menstrual pain were discovered through a rigorous protocol [50]:
- Participant Recruitment & Phenotyping: 20 women with regular cycles were stratified into mild (NPRS<4, n=12) and severe (NPRS≥4, n=8) pain groups based on numerical pain rating scale scores.
- Minimally-Invasive Sampling: Collected approximately 100 µL of finger-prick blood across menstrual, follicular, and luteal phases to minimize participant burden and enable dense longitudinal sampling.
- Multi-Platform Metabolomics: Employed five complementary analytical methods to quantify 692 metabolites encompassing steroid hormones (23), lipid mediators (85), abundant lipids (429), and hydrophilic metabolites (155).
- Data Integration & Validation: Used partial least squares-discriminant analysis (PLS-DA) to identify discriminatory metabolites and receiver operating characteristic (ROC) analysis to validate diagnostic performance of biomarker ratios.
Sebum Lipidomics Across the Menstrual Cycle
A 2025 study investigating sebum composition variations employed the following detailed methodology [32]:
- Cohort Design: 38 Chinese women aged 24-29 were categorized into High Sebum (HS, ≥150 µg/cm²) and Low Sebum (LS, ≤70 µg/cm²) groups using a Sebumeter.
- Standardized Sampling: Participants washed faces with a standardized cleanser, followed by isopropanol wipe to remove surface sebum. Sebum was collected using Cuderm S100 Sebutape patches applied to forehead and cheeks for 30 minutes during ovulation phase (OP) and early luteal phase (ELP).
- Lipid Extraction & Analysis: Sebum lipids were extracted with ethanol containing 0.025% BHT (antioxidant). Lipid classes (triglycerides, wax esters, squalene, free fatty acids) were quantified via GC-MS and LC-MS.
- Phase Determination: Menstrual cycle phases were determined by participant-recorded bleeding days and ovulation tests.
Large-Scale Metabolic Phenotyping
For population-level studies like the UK Biobank analysis [76]:
- Sample Processing: Serum and plasma were collected, centrifuged twice at 2500 × g for 15 minutes, aliquoted, snap-frozen in liquid nitrogen, and stored at -80°C.
- NMR Spectroscopy: Serum samples were diluted 1:1 with sodium phosphate buffer and analyzed using a Bruker Avance Neo 600 MHz spectrometer for absolute quantification of 168 metabolic markers.
- Statistical Modeling: A deep residual multitask neural network was trained on metabolomic markers to simultaneously learn disease-specific metabolomic states for 24 conditions, validated in four independent cohorts.
Signaling Pathways and Metabolic Relationships
The connection between metabolic signatures and clinical endpoints operates through several biological pathways that can be visualized to understand mechanism.
Modifying Factors and Confounders
The relationship between metabolic signatures and clinical endpoints is not uniform across populations but is significantly modified by various factors.
Menstrual Cycle Phase as a Metabolic Modifier
Cholesterol profiles exhibit a non-linear relationship with menstrual cycle phase. Analysis of 8,694 regularly menstruating women in UK Biobank revealed significant variations in total (p<0.001), HDL (p<0.001), and LDL (p=0.012) cholesterol across the cycle, though not in glucose or triglycerides [61]. These associations were moderated by body composition and activity levels, with greater metabolic variation across the cycle observed in the highest and lowest quartiles of fat mass and physical activity, respectively [61].
Nutritional and Lifestyle Modifiers
A cross-sectional study of 321 Japanese women identified specific nutritional factors associated with menstrual pain severity [77]:
- Protective Nutrients: Higher intake of animal proteins, vitamin D, and vitamin B12 was significantly associated with reduced pain severity (p<0.05).
- Lifestyle Factors: Regular breakfast consumption and bathing frequency were protective practices, while PMS symptom rates were significantly higher in the severe pain group (p<0.05).
The Scientist's Toolkit: Essential Research Reagents
Implementing these methodologies requires specific reagents and analytical tools optimized for metabolic profiling in menstrual cycle research.
Table 3: Essential Research Reagents for Metabolic Signature Studies
| Reagent/Instrument | Primary Function | Example Application | Technical Notes |
|---|---|---|---|
| Cuderm S100 Sebutape | Sebum collection via adhesive patches | Non-invasive sebum lipid sampling [32] | Patches applied 30min; store at -80°C |
| GC-MS / LC-MS Systems | Quantification of lipid classes & metabolites | Analysis of triglycerides, WEs, squalene, FFAs [32] | Use BSTFA+1% TCMS for derivatization |
| 600 MHz NMR Spectrometer | Absolute quantification of serum metabolites | Population-level metabolomic profiling [76] | Dilute 1:1 with phosphate buffer |
| Tanita BC-418 MA | Body composition analysis | Measure fat mass, muscle mass [77] | Account for clothing weight (set to 500g) |
| Sebumeter SM 815 | Sebum secretion rate measurement | Categorize High/Low Sebum groups [32] | Threshold: ≤70 μg/cm² (LS), ≥150 μg/cm² (HS) |
| Internal Standard Mix | Quantification standardization | d6-Cholesterol, d6-Squalene, d17C18:1 [32] | Include antioxidant (0.025% BHT) in extraction |
The integration of dynamic metabolic profiling with clinical endpoint assessment represents a paradigm shift in objective pain measurement and performance prediction. The rhythmicity of amino acids and lipids across the menstrual cycle provides a biological framework for understanding temporal patterns in symptom severity and functional capacity. Specific metabolite pairs, particularly the ratio of phosphatidylinositols to branched-chain amino acids, demonstrate exceptional discriminatory power for pain severity assessment. For researchers and drug development professionals, these advances offer new avenues for objective endpoint measurement in clinical trials, patient stratification strategies, and targeted therapeutic interventions that account for the inherent metabolic rhythmicity of the human body.
Benchmarking Against Established Clinical Chemistries and Hormone Profiles
Within the expanding field of female physiology research, the investigation of amino acid and lipid rhythmicity across the menstrual cycle represents a frontier for understanding metabolic health and developing novel interventions. A critical component of this research is the rigorous benchmarking of new metabolomic findings, such as oscillating amino acid and lipid species, against established clinical chemistries and hormone profiles. This practice anchors novel discoveries to a framework of well-understood physiological processes, ensuring that observed fluctuations are contextualized within the definitive hormonal milestones of the cycle. This technical guide outlines the protocols and analytical frameworks for integrating traditional endocrine measurements with advanced metabolomic profiling, providing researchers and drug development professionals with the methodology to build a validated and clinically relevant evidence base.
Core Hormonal and Metabolic Rhythmicity in the Menstrual Cycle
The menstrual cycle is characterized by predictable fluctuations in key reproductive hormones, which can be partitioned into several phases. For research purposes, a common model uses five phases: Menstrual (M), Follicular (F), Periovulatory (O), Luteal (L), and Premenstrual (P) [8]. The primary hormones of interest are estradiol (E2), progesterone (P4), luteinizing hormone (LH), and follicle-stimulating hormone (FSH).
These hormonal shifts govern the systemic metabolic environment. A foundational 2018 metabolomics study demonstrated that of 397 metabolites tested, 208 changed significantly (p < 0.05) across the cycle, with 71 meeting a False Discovery Rate (FDR) threshold of 0.20 [8] [7]. This rhythmicity was particularly pronounced in amino acids and their derivatives, lipid species, neurotransmitter precursors, and compounds involved in glutathione metabolism and the urea cycle [8]. The most significant finding was a widespread decrease in amino acids and specific lipids during the luteal phase, suggesting a state of increased nutrient utilization or anabolic demand [8].
Table 1: Characteristic Hormone and Metabolite Patterns Across the Menstrual Cycle
| Cycle Phase | Representative Hormone Profile | Characteristic Metabolic Signature |
|---|---|---|
| Menstrual (M) | Low E2, Low P4 | Recovery of amino acid and lipid levels; 39 amino acids and derivatives and 18 lipid species were lower in the preceding luteal phase [8]. |
| Follicular (F) | Rising E2, Low P4 | Steady-state or increasing amino acid levels. |
| Periovulatory (O) | LH/FSH Surge, High E2 | Transition period for metabolites. |
| Luteal (L) | High P4, High E2 | Significant decrease in plasma amino acids and specific lipid species [8]. |
| Premenstrual (P) | Falling E2, Falling P4 | Beginning of metabolite recovery. |
Methodologies for Phase Verification and Metabolite Quantification
Gold-Standard Hormonal Phase Verification
Accurate phase classification is paramount. Relying on self-reported cycle day counting alone is insufficient due to high inter-individual variability and the prevalence of subtle menstrual disturbances like anovulatory or luteal phase deficient cycles, which can occur in up to 66% of exercising females [48]. The gold standard requires direct hormonal measurement.
- Serum Hormone Assays: The highest level of accuracy for E2, P4, LH, and FSH is provided by serum immunoassays. Luteal phase verification should confirm a sufficient progesterone rise, with testing recommended 7–9 days post-LH surge detection to ensure ovulation and exclude anovulatory cycles [49].
- Urinary Luteinizing Hormone: Urinary LH surge detection is a practical and valid method for pinpointing the periovulatory phase and anchoring subsequent phase timing [8] [78].
- Salivary Hormone Testing: While feasible and non-invasive, salivary assays for E2 and P4 have shown inconsistencies in validity and precision in the literature. Their use requires careful validation against serum standards [78].
Table 2: Essential Research Reagent Solutions for Menstrual Cycle Research
| Reagent / Material | Function in Research | Technical Notes |
|---|---|---|
| Serum Immunoassay Kits (E2, P4, LH, FSH) | Gold-standard phase verification. | Critical for defining hormonally discrete phases and excluding participants with subtle menstrual disturbances. |
| Urinary LH Dipsticks | Non-invasive detection of the LH surge. | Used to forecast and confirm ovulation for timing periovulatory and luteal phase sampling. |
| LC-MS/MS Platform | Quantitative analysis of amino acids, lipids, and hydrophilic metabolites. | Provides high sensitivity and specificity for broad-spectrum metabolomic profiling [8] [50]. |
| GC-MS Platform | Complementary analysis of volatile metabolites and lipids. | Often used in tandem with LC-MS for comprehensive metabolomic coverage [8]. |
| HPLC-FLD System | Targeted analysis of specific micronutrients (e.g., B vitamins). | Used for specific analyte classes with high precision [8]. |
Advanced Metabolomic Profiling Techniques
- Sample Collection: Plasma is the preferred matrix for amino acid and lipidomic profiling. Fasting samples (minimum 4-8 hours) are required to minimize dietary confounding [79] [80]. Recent advancements allow for comprehensive analysis from minute volumes, such as 100 µL of finger-prick blood, facilitating frequent sampling across the cycle [50].
- Analytical Platforms:
- Liquid Chromatography-Mass Spectrometry (LC-MS) and Gas Chromatography-Mass Spectrometry (GC-MS) are workhorse platforms for untargeted and targeted metabolomics and lipidomics [8].
- High-Performance Liquid Chromatography with Fluorescence Detection (HPLC-FLD) is used for targeted nutrient analysis, such as B vitamins [8].
- Data Processing: Raw data undergoes peak identification, alignment, and normalization. Statistical analysis involves comparing phase means using appropriate mixed-effects models to account for within-participant repeated measures. Significance is evaluated against a p-value threshold (e.g., p < 0.05) with correction for multiple testing (e.g., FDR < 0.20) [8].
Benchmarking Metabolomic Data to Clinical Endpoints
The power of amino acid and lipid rhythmicity research lies in correlating these patterns with clinical hormone profiles and health outcomes.
Establishing Correlative Relationships
A 2018 study provides a prime example of benchmarking. It reported quantitative effect sizes for the contrast between the Luteal (L) and Follicular (F) phases for several amino acids, all with an FDR (q) < 0.20 [8]:
- Threonine: Effect Size = -0.45, q L-F = 6.73E-09
- Ornithine: Effect Size = -0.35, q L-F = 2.12E-05
- Arginine: Effect Size = -0.34, q L-F = 5.51E-04
- Alanine: Effect Size = -0.35, q L-F = 3.93E-04
These statistics quantitatively benchmark the significant decrease of these amino acids during the progesterone-dominant luteal phase against the established hormonal profile.
Linking Metabolites to Health Outcomes
A 2025 study on menstrual pain (dysmenorrhea) further demonstrates this principle. The research identified Branched-Chain Amino Acids (BCAAs - valine, leucine, isoleucine) and the phospholipid phosphatidylinositol (36:2) as potential biomarkers. It found an inverse correlation where PI(36:2) was elevated and BCAAs were reduced in a severe pain group [50]. The ratio of PI(36:2) to total BCAAs showed an exceptionally high accuracy for discriminating pain severity (AUC = 0.990) and correlated positively with pain scores, effectively benchmarking a novel metabolic signature against a clinically relevant subjective outcome [50].
Experimental Workflow and Metabolic Pathways
The following diagram illustrates the integrated experimental workflow for generating and benchmarking metabolomic data in menstrual cycle research.
Metabolic Pathways of Interest: The rhythmic metabolites identified in key studies participate in critical biochemical pathways. Fluctuations in ornithine, citrulline, and arginine point to a rhythmicity in the urea cycle [8]. Similarly, changes in glutamine and glycine suggest alterations in glutathione metabolism, indicating cyclic shifts in antioxidant capacity [8]. The involvement of amino acids as precursors for neurotransmitters (e.g., tryptophan for serotonin) provides a potential biochemical basis for mood and behavioral changes associated with the premenstrual phase [8].
The integration of robust hormonal phase verification with advanced metabolomics is non-negotiable for high-quality research into menstrual cycle rhythmicity. By systematically benchmarking novel findings of oscillating amino acids and lipids—such as the characteristic luteal phase decline—against the established framework of clinical chemistries and hormone profiles, researchers can move beyond mere observation to mechanistic insight. This rigorous approach provides the foundation for developing evidence-based, cycle-aware nutritional strategies and therapeutic interventions, ultimately advancing female health across the lifespan.
Conclusion
The rhythmic fluctuations of amino acids and lipids throughout the menstrual cycle represent a fundamental, yet historically overlooked, dimension of female physiology. This synthesis confirms a consistent pattern of metabolic downregulation, particularly for amino acids and specific phospholipids, during the luteal phase, likely supporting the anabolic demands of endometrial preparation. The emergence of robust biomarker ratios, such as phosphatidylinositol to branched-chain amino acids, demonstrates high diagnostic accuracy for conditions like dysmenorrhea, underscoring their potential in drug development and personalized medicine. Future research must prioritize standardized methodological frameworks, larger diverse cohorts, and the integration of multi-omics data to fully elucidate the molecular mechanisms driving these rhythms. Ultimately, incorporating menstrual cycle metabolic rhythmicity into research paradigms is not merely a technical consideration but a necessity for developing effective, sex-specific therapeutics and advancing precision health for women.
References
- 1. Menstrual cycle rhythmicity: metabolic patterns in healthy ... [https://www.nature.com/articles/s41598-018-32647-0]
- 2. Estrogen and Metabolism: Navigating Hormonal Transitions ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12431702/]
- 3. Hormonal regulation of metabolism—recent lessons learned ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10031253/]
- 4. Association between menstrual cycle phase and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10709933/]
- 5. Branched-chain amino acids and specific ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11762980/]
- 6. The central signaling pathways related to metabolism ... [https://translational-medicine.biomedcentral.com/articles/10.1186/s12967-025-06656-3]
- 7. Menstrual cycle rhythmicity: metabolic patterns in healthy ... [https://pubmed.ncbi.nlm.nih.gov/30275458/]
- 8. Menstrual cycle rhythmicity: metabolic patterns in healthy ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6167362/]
- 9. The influence of menstrual cycle on metabolic control and diet ... [https://ojrd.biomedcentral.com/articles/10.1186/s13023-025-03890-2]
- 10. The influence of menstrual cycle on metabolic control and ... [https://pubmed.ncbi.nlm.nih.gov/40653469/]
- 11. Effects of menstrual cycle phase on metabolomic profiles in ... [https://pubmed.ncbi.nlm.nih.gov/20150174/]
- 12. Leucine Requirements During the Menstrual Cycle [https://www.centerwatch.com/clinical-trials/listings/NCT06533501/leucine-requirements-during-the-menstrual-cycle]
- 13. Progesterone-associated arginine decline at luteal phase of ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0200489]
- 14. The influence of menstrual cycle on metabolic control and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12257650/]
- 15. MUSCLE PROTEIN METABOLISM AND ... [https://www.gssiweb.org/en/sports-science-exchange/article/muscle-protein-metabolism-and-protein-requirements-for-female-athletes--aligning-science-with-sex-specific-needs]
- 16. Cyclic Change of Fatty Acid Composition in Meibum During ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6736406/]
- 17. Variations in lipid levels according to menstrual cycle phase [https://pmc.ncbi.nlm.nih.gov/articles/PMC3130301/]
- 18. Ovarian lipid metabolic alterations in polycystic ovary ... [https://www.frontiersin.org/journals/medicine/articles/10.3389/fmed.2025.1652954/full]
- 19. The impact of menstrual cycle phase and symptoms on ... [https://www.frontiersin.org/journals/physiology/articles/10.3389/fphys.2025.1663657/full]
- 20. Quantitative Comparison of Statistical Methods for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9227835/]
- 21. Strategies for Comparing Metabolic Profiles [https://pmc.ncbi.nlm.nih.gov/articles/PMC5708160/]
- 22. Menstrual Cycle and Protein Requirements of Women [https://www.sciencedirect.com/science/article/abs/pii/S0022316623197288]
- 23. Warburg-like Metabolic Reprogramming in Endometriosis [https://pmc.ncbi.nlm.nih.gov/articles/PMC12195805/]
- 24. Molecular Impact of Metabolic and Endocrine Disturbance on ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12563804/]
- 25. Molecular Impact of Metabolic and Endocrine Disturbance ... [https://www.preprints.org/manuscript/202509.1235/v1]
- 26. A big-data approach to understanding metabolic rate and ... [https://elifesciences.org/articles/53560]
- 27. The strategic role of lipidomics in biomarker identification ... [https://www.frontiersin.org/journals/endocrinology/articles/10.3389/fendo.2025.1546512/full]
- 28. Metabolome analysis as a potential source of ... [https://www.nature.com/articles/s41598-025-22598-8]
- 29. Summary of Study ST003984 [https://www.metabolomicsworkbench.org/data/DRCCMetadata.php?Mode=Study&StudyID=ST003984&StudyType=&ResultType=]
- 30. 2025: new menstrual health research | Avon Longitudinal ... [https://www.bristol.ac.uk/alspac/news/2025/new-menstrual-health-research.html]
- 31. Machine learning-based menstrual phase identification ... [https://www.nature.com/articles/s44294-025-00078-8]
- 32. Physiological Differences in Sebum Composition in Regularly ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12592595/]
- 33. Mapping the temporal and spatial dynamics of the human ... [https://www.nature.com/articles/s41588-021-00972-2]
- 34. Mapping changes in women's visual functions during the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9634831/]
- 35. Natural menstrual rhythm and oral contraception diversely ... [https://www.nature.com/articles/s41598-018-29221-z]
- 36. Machine learning model for menstrual cycle phase ... [https://www.sciencedirect.com/science/article/pii/S0010482525000551]
- 37. Metabolic profiling of charged metabolites in association ... [https://www.sciencedirect.com/science/article/pii/S0378512221002954]
- 38. Evaluating finger-prick blood collection for remote ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12245952/]
- 39. how fingerstick blood testing is changing healthcare [https://www.neoteryx.com/microsampling-blog/how-fingerstick-blood-testing-is-changing-healthcare]
- 40. Stanford Medicine researchers measure thousands of ... [https://med.stanford.edu/news/all-news/2023/01/blood-drop-microsample-snyder.html]
- 41. Finger Stick Sampling Efficient Techniques: Learn How Now [https://www.diamonddiagnostics.com/blog/finger-stick-sampling-efficient-techniques]
- 42. Effect of Menstrual Cycle Phases on Plasma Lipid and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5483655/]
- 43. - Multi , Interpretation, and Its Application - PMC omics Data Integration [https://pmc.ncbi.nlm.nih.gov/articles/PMC7003173/]
- 44. Pathway enrichment analysis and visualization of omics data ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6607905/]
- 45. Pathway enrichment analysis and visualization of omics ... [https://cytoscape.org/cytoscape-tutorials/protocols/enrichmentmap-pipeline/]
- 46. Visualize · Pathway Guide [https://www.pathwaycommons.org/guide/workflows/rna_seq_to_enrichment_map/visualize/]
- 47. Unveiling Amino Acid Analysis: Challenges and Best ... [https://dmpkservice.wuxiapptec.com/articles/458-unveiling-amino-acid-analysis-challenges-and-best-practices-with-biological-samples/]
- 48. Why We Must Stop Assuming and Estimating Menstrual ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12152053/]
- 49. Menstrual Cycle Tracking in Sports Research [https://www.mdpi.com/2673-9488/3/4/44]
- 50. Branched-chain amino acids and specific ... [https://www.nature.com/articles/s41598-025-87415-8]
- 51. Establishing a Gold Standard for Quantitative Menstrual ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10533176/]
- 52. Higher central circadian temperature amplitude is ... [https://www.nature.com/articles/s41598-024-67297-y]
- 53. The Interplay between Early Mealtime, Circadian Rhythms ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10528427/]
- 54. Dietary circadian rhythms and cardiovascular disease risk ... [https://www.nature.com/articles/s41467-023-43444-3]
- 55. Association of diet, lifestyle, and chronotype with metabolic ... [https://www.nature.com/articles/s41598-024-55715-0]
- 56. Circadian Syndrome Is Associated with Dietary Patterns ... [https://www.mdpi.com/2072-6643/16/6/760]
- 57. Circadian rhythm phase shifts caused by timed exercise ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7098792/]
- 58. noisyR: enhancing biological signal in sequencing ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8373073/]
- 59. Untargeted Metabolomics: 5 Problems, 1 Solution [https://www.elucidata.io/blog/untargeted-metabolomics-5-problems-1-solution]
- 60. Menstrual cycle rhythmicity: metabolic patterns in healthy ... [https://hero.epa.gov/hero/index.cfm/reference/details/reference_id/7389501]
- 61. Association between menstrual cycle phase and metabolites ... [https://bmcmedicine.biomedcentral.com/articles/10.1186/s12916-023-03195-w]
- 62. Individual metabolomic signatures of circadian misalignment ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6581237/]
- 63. Genetics-nutrition interactions control diurnal enhancer ... [https://www.sciencedirect.com/science/article/abs/pii/S1550413125003560]
- 64. The microbiome interacts with the circadian clock and ... [https://elifesciences.org/articles/97130]
- 65. Association of Plasma Branched Chain Amino Acid with ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8496994/]
- 66. Using Phosphatidylinositol Phosphorylation as Markers for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7177416/]
- 67. Comprehensive bioinformatics and immunohistochemical ... [https://www.nature.com/articles/s41598-025-97118-9]
- 68. Branched-chain amino acids and specific ... [https://pubmed.ncbi.nlm.nih.gov/39863658/]
- 69. BCAA metabolism: the Achilles' heel of ovarian cancer ... [https://www.frontiersin.org/journals/endocrinology/articles/10.3389/fendo.2025.1579477/full]
- 70. Comparing the metabolomic landscape of polycystic ovary ... [https://www.nature.com/articles/s43856-025-00985-6]
- 71. A Comparative Analysis of PCOS, POR, and NOR - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC12271243/]
- 72. Visual analysis of multi-omics data [https://pubmed.ncbi.nlm.nih.gov/39318761/]
- 73. Association between menstrual phase and metabolites in... cycle [https://biomedcentral.eu/articles/10.1186/s12916-023-03195-w]
- 74. Frontiers | Crosstalk between dysregulated amino sensing and... acid [https://www.frontiersin.org/journals/oncology/articles/10.3389/fonc.2025.1665056/full]
- 75. Transcriptional programming of lipid and amino ... | PLOS Biology acid [https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.2005886]
- 76. Metabolomic profiles predict individual multidisease ... [https://www.nature.com/articles/s41591-022-01980-3]
- 77. of Severity Is Associated with Nutritional Intake and... Menstrual Pain [https://pmc.ncbi.nlm.nih.gov/articles/PMC10178419/]
- 78. Complexities in salivary and urinary methodology for ... [https://gremjournal.com/journal/01-2025/complexities-in-salivary-and-urinary-methodology-for-ovulation-and-menstrual-cycle-hormone-detection-a-scoping-review/]
- 79. AAp Amino Acids Profile | Amino Acids Test [https://www.diagnosticsolutionslab.com/tests/amino-acids-profile]
- 80. Amino Acid Analysis Plasma Test [https://www.ultalabtests.com/test/amino-acid-analysis-lc-ms-plasma?srsltid=AfmBOooqKWvFTRxuuIQc6FCpoMjU4heM5DexTx44OzEPDkriKcTVMKcM]